|
Evaluación In vitro de la producción de metano en variedades de pastos neozelandeses del altiplano de Nariño
ARTÍCULO DE INVESTIGACIÓN
José Edmundo Apráez-Guerrero
1Investigador, Facultad de Ciencias Pecuarias, Programa de Zootecnia, Universidad de Nariño. 2Programa de Zootecnia, Universidad de Nariño
This email address is being protected from spambots. You need JavaScript enabled to view it.
(Recibido: 25 de Mayo de 2016 Aprobado: 18 de Noviembre de 2016 Actualizado: 15 de diciembre de 2016)
DOI: 10.17151/vetzo.2016.10.2.8
RESUMEN: El calentamiento global es resultante de las actividades antrópicas, que producen altas cantidades de gases efecto invernadero (CO2, NH3, CH4), donde el metano posee un efecto mayor al dióxido de carbono y en el que la ganadería bovina es la principal causante de su emisión, como consecuencia de la fermentación anaerobia que sucede en el rumen. Las ganaderías especializadas del trópico alto de Nariño han masificado el uso de gramíneas neozelandesas en la alimentación de sus hatos, sin que hasta ahora se haya evaluado la producción de gases en esos pastos, hecho que motivó esta investigación en los forrajes: sterling (T1); nui (T2); ohau (T3) y samson (T4), mediante el uso de la técnica in vitro de producción de gases, utilizando fuente de inóculo fecal. T4 presentó mayor (P<0,05) producción de metano (37,26 ml CH4/g MSI), respecto a T1 (30,91), T2 (25,62) y T3 (25,51); sin embargo, la cantidad total del gas fue inferior (P<0,05) en T1 (0,45 ml CH4/ml GP), con relación a T2 (0,56), T3 (0,56) y T4 (0,99). La producción de metano se correlacionó positivamente (P<0,05) con la proteína verdadera (0,814) y negativamente (P<0,05) con FDN (-0,879) y FDA (-0,979). El ácido acético no fue detectado en T1 yT4, mientras que el butírico no se encontró en todos los tratamientos. La ecuación de regresión obtenida a través de lecturas de presión y volumen para T1 y T4 fue: Y= 4,9495 + 4,9519X - 0,0384X2 mientras que para T2 y T3 Y= 0,9812 + 7,5066X - 0,2715X2. El análisis fitoquímico no evidenció contenido de saponinas y un nivel moderado de fenoles en T1. No hubo diferencias (P<0,05) en la degradabilidad de MS, cuyos valores fueron: 56,48%, 59,37%, 49,37% y 58,25%, para T1, T2, T3 y T4 respectivamente.
Palabras clave: ácidos grasos volátiles, efecto invernadero, fermentación, medio ambiente, rumiantes
In Vitro evaluation of methane production in New Zealand pasture varieties of the high plateau of Nariño
ABSTRACT: Global warming resulting from human activities that produce high amounts of greenhouse gases (CO2, NH3, CH4) where methane has a greater effect than carbon dioxide and where cattle farming is the main cause of its emission as a consequence of the anaerobic fermentation that happens in the rumen. Specialized stockbreeding farms in the high Tropic of Nariño have made accessible to all the use of New Zealand grasses in the feeding of their herds, without having evaluated gas production in these grasses, a fact that motivated this research in pastures such as sterling (T1); nui (T2); ohau (T3) and samson (T4), through in vitro gas production technique using fecal inoculum source. T4 showed higher (P<0.05) production of methane (CH4 37.26 ml/g MSI) compared to T1 (30.91), T2 (25.62) and T3 (25.51). However, the total amount of gas was lower (P<0.05) in T1 (0.45 ml CH4/ml GP), compared to T2 (0.56), T3 (0.56) and T4 (0.99). Methane production was positively correlated (P<0.05) with the true protein (0.814) and negatively correlated (P<0.05) with FDN (-0,.879) and FDA (-0.979). Acetic acid was not detected in T1 and T4, whereas the butyric acid was not found in all treatments. The regression equation obtained through pressure and volume readings for T1 and T4 was: Y = 4.9495 + 4.9519X - 0.0384X2 while for T2 and T3 was Y = 0.9812 + 7.5066X - 0.2715X2. The phytochemical analysis did not evidence saponin content and a moderate level of phenols in T1. There were no differences (P<0.05) in the degradability of MS, whose values were 56.48%, 59.37%, 49.37% and 58.25% for T1, T2, T3 and T4 respectively.
Key words: environment, volatile fatty acids, fermentation, greenhouse effect, ruminants
Introducción
El inventario bovino en Colombia, según el ICA (2015), es de 22.527.783 animales, de los cuales 373.421 corresponden al departamento de Nariño y 34.702 para el municipio de Pasto, convirtiéndose esta actividad en una de las más relevantes dentro del sector pecuario de la región. Los rumiantes se consideran grandes contribuyentes al calentamiento global y deterioro de la capa de ozono, debido a la liberación de altas cantidades de dióxido de carbono y metano a la atmosfera (Carmona et al., 2005). Según Steinfeld (2009), la fermentación entérica y la descomposición del estiércol generan cerca del 80% de las emisiones de metano procedentes de la agricultura y aproximadamente el 35-40% del total de las emisiones antropogénicas, con un efecto contaminante 21 veces mayor que el CO2.
La técnica in vitro desprecia factores inherentes al animal como la absorción de acidos grasos volátiles (AGV), liberación de amoniaco desde compuestos nitrogenados (Van Lier & Regueiro, 2008) y tasa de pasaje, los cuales pueden causar variaciones en el pH, formación de AGV, crecimiento microbiano y producción de metano (Noguera et al., 2006; Knapp et al., 2014), limitando la comparación in vivo. Los bovinos disponen de un sistema digestivo capaz de aprovechar y convertir material fibroso en alimentos de alta calidad nutritiva; sin embargo, por sus características innatas producen CH4, CO2 y AGV (Montenegro & Abarca, 2002), la formación de CH4 es el principal modo de eliminación de hidrógeno generado como producto principal de la fermentación microbiana (Moss et al., 2000), responsable por la pérdida de energía aportada por el alimento (Carmona et al., 2005).
Además de poseer elevada palatabilidad y digestibilidad, floración precoz a intermedia, alta producción de macollos, entre otras características, los pastos neozelandeses Lolium hibridum (sterling-ohau) y Lolium perenne (nui-samson), presentan el endófito AR1, hongo que otorga mayor persistencia y resistencia a la sequías y protege la planta contra el ataque de insectos como el Listronotus bonariensis (Demanet, 2008). Según MEGALAC (2009), el área sembrada con estos pastos en Nariño es aproximadamente 10.103 hectáreas, de las cuales el 20,22% corresponden a raigrases (Lolium perenne). El objetivo del presente estudio fue evaluar la producción de metano de las principales variedades de pasturas neozelandesas utilizadas en la ganadería de la región mediante la técnica in vitro, apoyándose en análisis nutricionales, complemento estadístico, además de la creación de una ecuación de regresión que permita estimar el volumen de gas generado acorde a la altura de los laboratorios de la Universidad de Nariño.
Materiales y Métodos
Localización
La investigación se desarrolló en las fincas La Frisona y El Encanto, ubicadas en los corregimientos de Gualmatan y Catambuco, respectivamente, con una temperatura promedio de 11ºC, altura de 2.800 msnm; precipitación pluvial anual de 703 mm y a 6 km del municipio de San Juan de Pasto, Colombia. De acuerdo con Holdridge (1998) corresponden a la zona de vida bosque seco Montano Bajo (bs-MB).
Preparación de muestras
Se evaluaron los tratamientos: T1 (sterling), T2 (nui), T3 (ohau) y T4 (samson); con cuatro repeticiones cada uno. Se colectaron las muestras en cada una de las praderas por medio de un marco de madera de 1 m2 (sierra apud Narváez & Delgado, 2011); posteriormente fueron secadas a 65°C durante 68 h hasta alcanzar peso constante y molidas a través de un molino estacionario marca Beb Mühlenbau Nossen modelo 8255 con una criba de 1 mm, al finalizar las muestras se depositaron en recipientes herméticos para su conservación y posterior análisis.
Análisis químico
Una muestra de cada sustrato fue llevada a los laboratorios especializados de la Universidad de Nariño para hacer los respectivos análisis según las metodologías planteadas por la AOAC (1995), el contenido de humedad y ceniza se determinó por medio del método termo gravimétrico, la grasa por el método Soxhlet, la proteína por Kjeldahl (Nx6.25), los minerales Ca, P y Mg por digestión húmeda espectrofotométrica, la fibra detergente acida, neutro y lignina por Van Soest, saponinas y fenoles por pruebas cualitativas.
Preparación de medio de digestión
Un día antes de iniciar la prueba, se preparó el medio de digestión para 30 botellas de 110 ml de capacidad siguiendo las recomendaciones de Narváez & Delgado (2011): 504,3 ml de agua destilada a la que se adicionó 1,02 g de caseína y 0,51 ml de solución micro mineral, 1020 ml de solución macro mineral, 1020 ml de solución buffer, 5,1 ml de resazurina, esta solución fue agitada para obtener una mezcla homogénea y saturada con CO2, por dos horas hasta que adquirió una leve coloración rosa.
Solución micromineral. Se disolvió en 3 ml de agua destilada precalentada a 39oC; 0,4 g cloruro de calcio dihidratado (CaCl2-2H2O); 0,3 g de cloruro de manganeso tetrahidratado (MnCl2-4H2O); 0,03 g de cloruro de cobalto hexahidratado (CoCl2-6H2O) y 0,24 g de cloruro férrico hexahidratado (FeCl3-6H2O).
Solución buffer. Se disolvió en 1020 ml de agua destilada y precalentada a 39oC; 4,08g de carbonato acido de amonio (NH4HCO3) y 35,7g de carbonato acido de sodio (NaHCO3).
Solución macromineral. Se disolvió en 1020 ml de agua destilada precalentada a 39oC; 4,56 g de ortofosfato acido disodico (Na2HPO4-12H2O); 6,32 g de dihidrogenofosfato de potasio (KH2PO4) y 0,612 g de sulfato de magnesio (MgSO4-7H2O).
Solución indicadora. Se disolvió 0,0075 g de (C12H6NO4Na) o rezasurina en 7.5 ml de agua.
Agente reductor. Se disolvió en 120 ml de agua, 0,75 g de cisteína HCl-HO2CCH (NH2) CH2SH, 4,8 g de Hidróxido de Sodio 1M (NaOH) 1M y 0,75 g de sulfato de sodio (Na2SO4).
Preparación de las botellas de incubación
Para que las botellas mantengan una temperatura constante de 39°C se adecuó como baño maría un acuario de 1 m de largo, 0,35 m de ancho y 0,35 m de alto, recubierto totalmente por una lámina de icopor de 1,2 cm de espesor junto con un termostato, evitando así fluctuaciones que provoquen variaciones de la temperatura y por ende en los resultados.
Se trabajó con un total de 26 botellas de incubación, correspondientes a 4 tratamientos con 4 repeticiones cada una, 2 blancos, los cuales contenían medio de digestión e inoculo fecal pero no sustrato, con el fin de obtener el total de gas realmente derivado desde la fermentación del sustrato (Posada & Noguera, 2005) y 2 estándares por duplicado conformados por dos mezclas: (aubade, samson, sterling, trébol rojo) y (saboya, aubade, samson, trébol rojo, diente de león) obtenidos de la investigación de Francisco Achicanoy. Además, cuatro botellas adicionales para la determinación de metano.
Las botellas antes del inicio de la prueba fueron lavadas y secadas en estufa a 105oC por 12 h y saturadas con CO2 para luego rotularlas con la información correspondiente.
Un día antes de la prueba, 1 g de sustrato fue llevado dentro de cada botella y utilizando una jeringa graduada se depositaron manualmente 85 ml de medio de digestión y 4 ml de solución reductora, las botellas fueron selladas con tapas de caucho, mediante un crimper manual; al terminar fueron refrigeradas a 4ºC para evitar alguna fermentación. Antes del inicio de la prueba las botellas fueron llevadas a una estufa de ventilación forzada a 39ºC.
Colección de inóculo
Los animales donantes fueron tres vacas Holstein de alto mestizaje con un peso promedio de 530 kg, de la Corporación Colombiana de Investigación Agropecuaria (CORPOICA), ubicada a 5 km de San Juan de Pasto, vía Pasto-Obonuco.
La dieta de los donantes consistió principalmente en pasto sterling de 30 días de edad promedio, suministrado durante ocho días anteriores a la colecta de las heces, las cuales fueron extraídas el día del inicio del experimento en horas de la mañana, mediante la técnica per rectum, depositadas en una garrafa térmica precalentada a 39°C (Narváez & Delgado, 2011), e inmediatamente transportados a los laboratorios de la Universidad de Nariño. Las heces fueron filtradas a través de un paño de gasa de cuatro capas, el material filtrado fue transferido a baño maría a 39ºC y continuamente saturado con CO2.
Cada botella se inoculó con 10 ml, se selló, agitó y trasfirió al baño maría, utilizando el transductor de presión se ajustó la presión interna a cero.
Medición de presión y volumen
Se utilizó un sistema a base de una válvula de tres salidas sobre las cuales se acopló una aguja de 0,7 mm, la cual fue insertada a través de cada una de las tapas de caucho que sellaban las botellas, se insertó una jeringa graduada la cual fue utilizada para medir el volumen de los gases acumulados en la parte superior de los frascos en las horas 0, 3, 6,12, 24, 48 y 72; la presión se registró por medio un transductor de presiones marca Autonics modelo PSA-1 con lector digital.
Al finalizar las lecturas de volumen y presión, las botellas fueron manualmente agitadas y reubicadas en el baño maría.
Determinación de metano
El gas se succionó con una jeringa y depositó en viales N20 de 10 ml, marca Peldar en vidrio color ámbar, previamente lavados y secados en estufa a 105°C por 12 h, los cuales fueron rotulados, sellados con corchos plásticos y ajustados su presión a cero. Una vez depositado, el gas se llevó inmediatamente a los laboratorios especializados de la Universidad de Nariño, para su posterior análisis por cromatografía (López & Newbold, 2007). El gas producido se extrajo de los viales con una jeringa (Headspace Soil gas SUPELCO) para cromatografía, e inyectado en un cromatógrafo (Modelo GC 17ª, marca SHIMADZU. Tokio, Japón), se tomaron 5 ml de muestra de los cuales se inyectaron 0,5. Se empleó una columna apolar (DB-5MS J&W Scientific, 30 m*0,25 mm*0,25 µm). Las temperaturas del puerto de inyección Split/Splieless y detector FID fueron 200°C y 250°C respectivamente, se realizó una programación de temperatura de la columna de 30°C /5 min hasta 200°C a reacción de 30°C/min. Como gas de arrastre se empleó Helio UAP a flujo de columna de 1,0 ml/min. El tiempo de retención del metano fue de 1,80 min. El software utilizado para la adquisición y el análisis de la información fue el CLASS VP versión 4.3.
Para determinar la degradabilidad de la materia seca, se utilizaron 20 crisoles previamente tarados y adecuados con fibra de vidrio para filtrar los residuos contenidos en cada botella de incubación, mediante un sistema de matraces con orificio de salida lateral y secados en estufa a 105°C El material degradado se calculó por diferencia de pesos entre los crisoles vacíos, el peso inicial de la muestra incubada y los crisoles con residuo tras el secado en estufa. (Narváez & Delgado, 2011)
Para obtener la ecuación de regresión se ingresaron los datos de presión y volumen registrados en las 0, 3, 6,12, 24, 48 y 72 h, al programa estadístico SAS 9.1; se obtuvo la gráfica y la correspondiente ecuación mediante el complemento PROC REG.
Se realizaron las correlaciones de metano con proteína cruda, proteína verdadera, FDA, FDN y materia seca, mediante el procedimiento CORR del paquete estadístico SAS 9.1.
Análisis estadístico
Los datos de volumen fueron analizados mediante el modelo de medidas repetidas en el tiempo con el complemento PROC MIXED SAS 9.1, correspondiente a cuatro tratamientos con cuatro repeticiones. Los datos de producción de metano y degradabilidad de materia seca fueron analizados mediante el diseño completamente al azar, utilizando PROC GLM SAS 9.1 y estudiando 4 tratamientos y 4 repeticiones; las medias fueron ajustadas y comparadas según la prueba de Tukey - Kramer (P<0,05). Para ello se encontró mejor ajuste con la estructura autorregresiva de primer orden, teniendo en cuenta el menor valor del criterio de información de Akaike.
Modelo matemático:
Resultados y Discusión
Análisis químico proximal
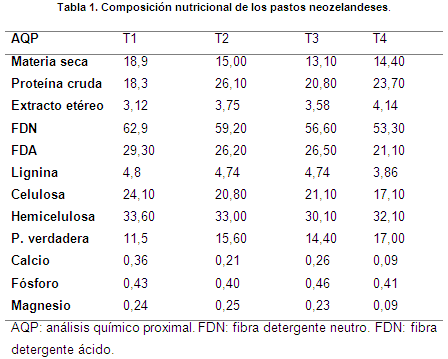
La composición nutricional de los pastos se consigna en la tabla 1. El contenido de materia seca de los tratamientos fue 15,35% en promedio, superior al obtenido por Posada et al. (2013) de 13,7% en raigrás híbrido tetraploide; por su parte, Achicanoy (2015) afirma que el contenido de materia seca es influenciado por los programas de fertilización utilizados, estado fenológico de la planta, manejo de la pradera, condiciones edafoclimáticas de cada finca y disponibilidad de agua.
El valor promedio de proteína cruda (PC) fue 22,23%, inferior al reportando por Villalobos & Sánchez (2010) de 25,21% para Lolium perenne. Al contrastar los pastos tetraploides con los perennes, se encuentra que los primeros tienen un porcentaje mayor de proteína, aunque su requerimiento en fertilización nitrogenada es más alta (Demanet, 2008); por consiguiente, se infiere que la cantidad de proteína es afectada si las pasturas se someten al mismo plan de fertilización
Los valores medios de FDN y FDA fueron 58% y 25,78%; respectivamente. Sin embargo, Carulla et al. (s.a) reportaron un contenido de FDN menor (47,4%) y FDA superior (28,6%). El promedio de hemicelulosa fue de 32,2%, superior al obtenido por Correa et al. (2008) de 26,2% para pasto kikuyo (Pennisetum clandestinum); según Herrera apud Fernández et al. (2014), estas variaciones en los carbohidratos estructurales pudieron ser causadas por la edad del pasto, ya que los componentes estructurales aumentan con la madurez, disminuyendo los carbohidratos solubles y afectando la calidad nutricional.
El promedio de lignina de los tratamientos fue 4,54%; un alto porcentaje de este constituyente implica el acceso limitado de los microorganismos del rumen a los nutrientes intracelulares, manifestando reducción en la digestibilidad (Villalobos, 2014).
El contenido de calcio fue 0,23% en promedio; su presencia en las plantas depende del manejo de las praderas, la edad de corte, el tiempo de permanencia de los animales o la época (Matute & Ayala, 2001). No obstante, el valor promedio reportado para el fosforo fue 0,43%; según Vásquez apud Ramírez et al. (2008) este mineral está presente en mayor proporción en brotes y hojas jóvenes y su cantidad refleja el estado fenológico de los forrajes. El porcentaje de magnesio de los tratamientos fue 0,20% en promedio, su bajo nivel puede ser ocasionado por la deficiente fertilización utilizada, ya que la concentración de este elemento es menor (Kirchner et al., 1990)
Producción de gas
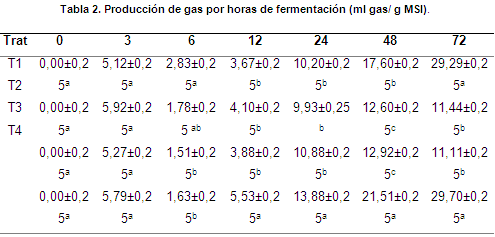
En la hora 3 no se observaron diferencias significativas entre los tratamientos (P>0,05) (tabla 2) pero sí en la hora 6 (P<0,05) (tabla 2). En la hora 3 observó un crecimiento en la producción de gas de todos los tratamientos (tabla 2), el cual decreció hasta la hora 6 en donde a su vez comienza a tomar una producción ascendente, pudiendo haber sido causado posiblemente por la fermentación de los carbohidratos solubles y la pectina, lo cual pudo ocasionar disminución en el pH y retardo en la fermentación de la celulosa debido a la inhibición de la flora celulolítica, cuando la mayor parte de estos carbohidratos se han fermentado, el pH probablemente subió y la degradación de la celulosa se reanudó (Van Lier & Regueiro, 2008).
A las 12 y 24 h, los tratamientos T1, T2 y T3 no presentaron diferencias significativas (P>0,05) entre ellos, pero sí respecto a T4 (P<0,05) (Tabla 2). En la hora 72 se evidenció que T1 y T4 presentaron diferencias significativas (P<0,05) respecto a T2 y T3; de la misma manera, la mayor formación de gas total fue presentada por T1 y T4 (tabla 2), al culminar las 72 h de fermentación se observó que T1 y T4 aún generaban elevada producción de gas, infiriendo que posiblemente su tiempo de fermentación puede ser mayor a 72 horas.
Ecuación de regresión relacionando presión y volumen de gas
Debido a las diferencias presentadas (P<0,05) en la generación de gas entre todos los tratamientos al culminar el periodo fermentativo de 72h, se realizaron dos ecuaciones de regresión, una que agrupa a T1 y T4 (y = 4,9495+4,9519x-0,038x2R2=0,98) y otra a T2 y T3 (y= 0,9812+7,5066x-0,2715x2 R2=0,98) obteniendo una relación presión: volumen de 1 psi =9,86ml y 1 psi=8,22 ml, respectivamente. Al compararlas con los resultados reportados por Narváez & Delgado (2011), quienes obtuvieron una ecuación de y = -0,0240 + 5,5118x + 0,0048x2 R2= 0,97, se evidenció mayor relación presión volumen, pese a que el inóculo utilizado fue fecal y las condiciones de altura sobre el nivel del mar del laboratorio eran iguales para los dos estudios, hubo diferencias en los animales donantes empleados, la dieta base, el número y variedad de muestras de sustratos, que probablemente pudieron ser las causantes de esta variación.
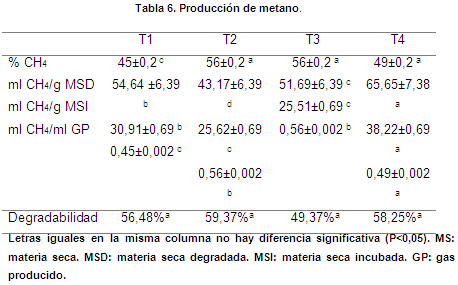
Producción de metano
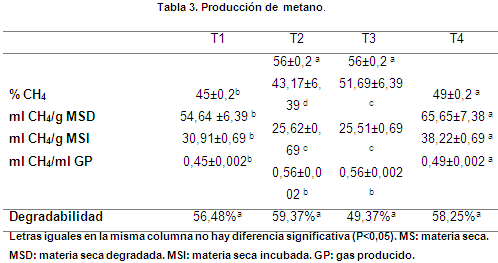
La producción de metano no presentó diferencias estadísticamente significativas (P>0,05) al comparar T2 (56%), T3 (56%) y T4 (49%); sin embargo, estos mismos tratamientos fueron diferentes (P<0,05) a T1 (45%) (Tabla 3); en cuanto a ml CH4/g materia seca degradada (MSD), hubo diferencias significativas (P<0,05), entre todos los tratamientos, existiendo una mayor producción de metano para T4 (65,65 ml CH4/g MSD). No obstante, no se reportaron diferencias significativas (P>0,05) entre T2 y T3 en ml CH4/g materia seca incubada (MSI), pero fueron diferentes a T1 y T4, siendo T4 el de mayor emisión (38,22 ml CH4/g MSI).
La presencia de metabolitos secundarios en las plantas son componentes de defensa contra ataques de los herbívoros, además tienen la capacidad de reducir las poblaciones bacterianas en el rumen y por ende las emisiones de metano (Puchala et al., apud Piñeiro, 2015), por lo tanto, se afirma que la baja cantidad de este gas en T1 (45%) pudo deberse a la presencia moderada de fenoles, pues existe una relación inversa entre el contenido de taninos en los forrajes con la producción de metano (Meale et al.,2012), como también el contenido de lignina, ya que dietas ricas en fibras lignificadas conducen a su menor producción, debido al acceso limitado de los microorganismos del rumen al sustrato (Kreuzer & Hindrichsen, 2006).
El uso de plantas con contenido tanífero en la alimentación del ganado lechero reducen las emisiones de metano y son una alternativa viable para su mitigación (Lascano & Cárdenas apud Santacoloma, 2011) Así mismo, la baja proporción de los componentes estructurales como la celulosa y hemicelulosa está asociada a una menor formación de metano entérico (Kreuzer & Hindrichsen, 2006), lo cual se evidencia en T4, cuya producción de CH4 es 49% y sus contenidos de FDA (21,1%) y FDN (53,3%) son inferiores en relación a los demás tratamientos.
La producción de CH4 en los rumiantes está influenciada por factores como el consumo y la digestibilidad del alimento, así como la composición de la dieta; desde este punto, se manifiesta que los forrajes de alta calidad reducen las emisiones de metano mediante la modificación de la fermentación ruminal (Bonilla & Lemus, 2012). De tal manera, que los tratamientos evaluados presentan variación en la producción de metano, ya que la calidad nutricional difiere entre los mismos.
Vargas et al. (2014) reportan un valor de 46,8 ml CH4/g MSD en una mezcla de raigrás y trébol 90:10, inferior a los obtenidos en esta investigación, a excepción de T2 (43,17 ml CH4/ g MSD) (Tabla 3). Los taninos en la leguminosa son la causa de que la formación de metano sea inferior en una mezcla con gramíneas, pues están inversamente relacionados con la emisión de este gas (Tekippe et al., 2013); Narváez & Delgado (2011) estimaron un valor de 37 ml CH4/g MSI en raigrás, superior a T1, T2 y T3.
Degradabilidad de la materia seca (DMS)
No hubo diferencias significativas (P>0,05) entre todos los tratamientos, infiriendo que la producción de gas no fue afectada por esta variable, puesto que mayor producción de gas no indica más eficiencia de los microorganismos en la utilización del sustrato, de tal forma que dos pastos con igual degradación de MS pueden originar diferentes volúmenes de gas (Noguera et al., 2006).
En esta investigación no se midió el coeficiente de partición que puede causar variación en los resultados al momento de comparar con otros estudios, el cual es generado por la variación que la biomasa microbiana puede causar en la relación sustrato verdaderamente degradado a volumen de gas producido (Posada & Noguera, 2005).
Correlaciones
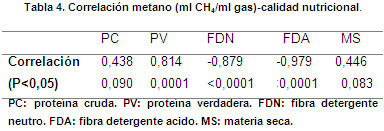
Se obtuvo correlación (P<0,05) negativa entre la FDN (-0,317) y FDA (-0,108) respecto a metano (tabla 4); por otro lado, Rivera et al. (2015) obtuvieron relación positiva entre CH4 con FDA y FDN, mientras que Patra & Lalhriatpuii (2016) reportaron correlación negativa con un grado bajo de significancia. El entramado tridimensional de los componentes y el nivel de lignificación de la pared celular son factores relacionados inversamente con el grado de adhesión de las bacterias fibrolíticas, tasa de fermentación y producción de metano, asociados al estado de madurez del forraje (De Blas et al., 2008; Calsamiglia, 1997).
Se observó correlación (P<0,05) positiva entre CH4 y contenido de proteína verdadera (0,462) (tabla 4); en el rumen la proteína microbiana corresponde al 80% de la proteína verdadera, existiendo una relación directa entre éstas, que generan cambio en la cantidad microbiana; por ende, en la colonización, fermentación y generación de gases (Mac, 2010).
Metabolitos secundarios
No hubo presencia de saponinas en los tratamientos (tabla 5), por tanto este no fue un factor que influyó en la producción de gas, aunque su presencia según Carmona et al. (2005) y Makkar apud Hristov et al. (2013), puede llegar a disminuir la producción de CH4 de 6 a 27% y el recuento de protozoarios que sin inhibir la actividad bacteriana, pueden causar la reducción en la digestibilidad de la materia orgánica
Los compuestos fenólicos son un amplio grupo de sustancias químicas, considerados metabolitos secundarios de las plantas; siendo los taninos parte de ellos (Martínez et al., 2000). El contenido fenólico fue moderado para T1 (tabla 5), causado posiblemente por la presencia de taninos, lo que evidencia el efecto negativo sobre la producción de metano (ml CH4/ ml gas). De acuerdo con Meale et al. (2012), el contenido de taninos en los forrajes está relacionado inversamente con la producción de metano no obstante, su presencia se asocia con la disminución de la digestibilidad de los nutrientes, particularmente de la proteína cruda.
Conclusiones
El porcentaje de metano generado por el pasto samson fue causado por su menor contenido de carbohidratos estructurales, convirtiéndolo en el menos contaminante de los pastos evaluados; por su alta producción de gas se considera una opción eficiente al aportar energía aprovechable para el animal. La presencia de fenoles en el pasto sterling redujo el porcentaje de metano, demostrando la acción inhibitoria que los metabolitos secundarios ejercen sobre su formación. Por el comportamiento demostrado de los pastos en la producción de volumen de gas respecto al tiempo de fermentación, se obtuvieron dos ecuaciones de regresión teniendo en cuenta la altitud de los laboratorios de la universidad de Nariño, una para los pastos samson y sterling y otra para nui y ohau, comprobado de esta manera que aunque todos estos fueron del genero Lolium, su comportamiento durante la técnica in vitro varió. Al igual que otros estudios, se evidenció que la producción de gas in vitro estuvo relacionada con la velocidad de fermentación de los carbohidratos, además del contenido de materia seca y proteína.
Referencias
Ahicanoy-Anganoy, F.J. Determinación de la emisión de gases efecto invernadero en dos sistemas productivos de la cuenca lechera de departamento de Nariño mediante el uso de la técnica in vitro. Pasto, Colombia: Universidad de Nariño, 2015. 115p. Tesis (maestría en ciencias agrarias).
Association of official analytical chemistry-AOAC. Official methods of analysis. 16 ed. Arlington: AOAC International, 1995. p. 2201-3301.
Bonilla-Cárdenas, J.A.; Lemus-Flores, C. Emisión de metano entérico por rumiantes y su contribución al calentamiento global y al cambio climático. Rev Mex Cienc Pecu, v.3, n.2, p.215-246, 2012.
Calsamiglia, S. Nuevas bases para la utilización de la fibra en dietas de rumiantes. Madrid, España: Universidad Autónoma de Barcelona, 1997. Disponible en: http://www.ucv.ve/fileadmin/user_upload/ facultad_agronomia/Uso_de_Fibra_en_Rumiantes.pdf. Accesado en: 30/04/2016.
Carmona, J.; Bolívar, D.; Giraldo, L. El gas metano en la producción ganadera y alternativas para medir sus emisiones y aminorar su impacto a nivel ambiental y productivo. Rev Col Cienc Pec, v.18, n.1, p.49-63, 2005.
Carulla, J.E.; Cárdenas, E.; Sánchez, N. et al. Valor nutricional de los forrajes más usados en los sistemas de producción lechera especializada de la zona andina colombiana, Bogotá: Universidad Nacional de Colombia; Disponible en: https://www.academia.edu/8158740/Valor_nutricional_ de_los_forrajes_en_colombia. Accesado en: 18/04/2016.
Correa, H.J.; Pabón, M.L.; Carulla, J.E. Valor nutricional del pasto kikuyo (Pennisetum clandestinum Hoechst Ex Chiov.) para la producción de leche en Colombia (Una revisión): I - Composición química y digestibilidad ruminal y posruminal. Livestock Research for Rural, v. 20, n. 4, p.1, 2008.
De Blas, C.; García-Rebollar, P.; Cambra-López, M. et al. Contribución de rumiantes a emisiones de gases con efecto invernadero: FEDNA, 2008. Disponible en: http://www.produccionbovina.com.ar/sustentabilidad/89-gases.pdf. Accesado en: 24/04/2016.
Demanet-Filippi, R. Manual de especies forrajeras y manejo de pastoreo. Programa Desarrollo Productores. Temuco, Chile: Universidad de la Frontera, 2008.
Fernández-Martínez, J.; Gómez-Angulo, I.; Cordoví -Castillo, E. et al. Rendimiento y composición bromatológica de Cynodon nlenfuensis cv. jamaicano en un suelo vertisol de la provincia Granma, Cuba. Revista Granma Ciencia, v.18, n.2, p.10, 2014.
Grupo de investigación producción y sanidad animal línea de genética y mejoramiento animal-MEGALAC. Caracterización y evaluación genética de la población bovina lechera del trópico alto de Nariño, para la conformación de núcleos de selección. Caracterización general de la población bovina lechera del trópico alto de Nariño, 2009.
Hristov, A.N.; Oh, J.; Firkins, J.L. et al. Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. Journal of Animal Science, v. 91, p.5045-5069, 2013.
Instituto Colombiano Agropecuario-ICA. Censo Pecuario Nacional: Censo Bovino en Colombia, 2015. Disponible en: http://www.ica.gov.co/getdoc/8232c0e5-be97-42bd- b07b-9cdbfb07fcac/Censos-2008.aspx. Accesado en: 15/03/2016.
Kirchner-Salinas, F.R.; Usami-Olmos, C.R.; López-González, E. Cultivos forrajeros. [S.I.]: Trillas, 1990.
Knapp, J.R.; Laur, G.L.; Vadas, P.A. et al. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. Journal Dairy Science, v. 97, p.3231-3261, 2014.
Kreuzer, M.; Hindrichsen, I.K. Methane mitigation in ruminants by dietary means: The role of their methane emission from manure. International Congress Series, v. 1293, p.199-208, 2006.
Kumar-Patra, A.; Lalhriatpuii, M. Development of statistical models for prediction of enteric methane emission from goats using nutrient composition and intake variables. Agriculture Ecosystems and Environment, v.215, p.89-99, 2016.
López, S.; Newbold, C.J. Analysis of methane. In: Makkar, H.P.; Vercoe, P.E. (Ed). Measuring Methane Production from Ruminants. Netherlands: Springer, 2007. p.1-13.
Mac-Loughlin, R.J. Requerimientos de proteína y formulación de raciones en bovinos para carne, 2010.http://www.produccionbovina.com/informacion_ tecnica/invernada_o_engorde_en_general/ 42-formulacion_proteina.pdf. Accesado en: 26/04/2016.
Martínez-Valverde, I.; Periago, M.J.; Ros, G. Significado nutricional de los compuestos fenólicos de la dieta. ALAN, v. 50, n. 1, p.1, 2000.
Matute-Estrada, G.M.; Ayala-Velásquez, P.S. Determinación de los niveles de minerales presentes en pastos y suero sanguíneo de vacas lecheras en el Litoral Atlántico de Honduras. Zamorano, Honduras: Carrera de Ciencia y Producción Agropecuaria, 2001. 31p. Tesis (pregrado ingeniería agronómica).
Meale, S.J.; Chaves, A.V; Baah, J. et al. Methane Production of Different Forages in in vitro Ruminal Fermentation. Asian-Australasian Journal of Animal Science, v. 25, n. 1, p.86-91, 2012.
Montenegro, J.; Abarca, S. Intensificación de la ganadería en Centroamérica: fijación de carbono, emisión de metano y de óxido nitroso en sistemas de producción bovina en costa rica. FAO, 2002. Disponible en: http://www.fao.org/documents/card/es/c/bc024e4e- 92fc-59e9-bb4b-cf8e629b15b0/. Accesado en: 16/03/2016.
Moss, A. Jouany, J.P.; Newbold, J. Methane production by ruminants: its contribution to global warming. Ann zootech., v.49, n.3, p.231-253,2000.
Narváez-Herrera, J.P.; Delgado-Rodríguez, J.M. Caracterización de recursos forrajeros herbáceos, arbóreos y arbustivos de uso convencional y alternativo en el trópico alto del departamento de Nariño, mediante el uso de la técnica in vitro de producción de gases. Pasto, Colombia: Universidad de Nariño, 2011.183p. Tesis (pregrado zootecnia).
Noguera, R.R.; Ramírez, C.I.; Bolívar, D.M. Efecto de la inclusión de papa (Solanum tuberosum) en la cinética de fermentación in vitro del pasto kikuyo (Pennisetum clandestinum). Livestock Research for Rural Development, v.18, n.5, p.1, 2006.
Piñeiro-Vázquez, A.T.; Canul-Solís, J.R.; Alayón-Gamboa, J.A. et al. Potencial de los taninos condensados para reducir las emisiones de metano entérico y sus efectos en producción de rumiantes. Arch Med Vet, v. 47, p. 263-272.
Posada, S.L.; Noguera, R.R. Técnica in vitro de producción de gases: Una herramienta para la evaluación de alimentos para rumiantes. Livestock Research for Rural Development, v.17, n.4, p.1, 2005.
Posada-Ochoa, S.; Cerón, J.M.; Arenas, J. et al. Evaluación del establecimiento de ryegrass (Lolium sp.) en potreros de kikuyo (Pennisetum clandestinum) usando la metodología de cero labranza. Revista CES Medicina veterinaria y Zootecnia, v.8, n.1, p.26-35, 2013.
Ramírez, J.L.; Verdecia, D.; Leonard, I. Rendimiento y caracterización química del Pennisetum Cuba CT 169 en un suelo pluvisol. Revista Electrónica de Veterinaria, v. 9, n.5, p.1-10, 2008.
Rivera, J.E.; Molina, I.C.; Donney`s, G. et al. Dinámica de fermentación y producción de metano en dietas de sistemas silvopastoriles intensivos con L. leucocephala y sistemas convencionales orientados a la producción de leche. Livestock Research for Rural Development, v. 27, n. 4, p.1-15, 2015.
Santacoloma-Varón, L.E. las dietas en las emisiones de metano durante el proceso de rumia en sistemas de producción bovina. Revista de investigación agraria y ambiental, v. 2, n. 1, p.55-64,2011.
Steinfeld, H.; Gerber, P.; Wassenaar, T. et al. La larga sombra del ganado problemas ambientales y opciones: el papel del ganado en el cambio climático y en la contaminación atmosférica. FAO, 2009. Disponible en: http://www.fao.org/3/a-a0701s.pdf. Accesado en: 15/03/2016.
Tekippe, J.A.; Tacoma, R.; Hristov, A.N. et al. Effect of essential oils on ruminal fermentation and lactation performance of dairy cows. Journal of Dairy Science, v. 96, p. 7892-7903, 2013.
Theodorou, M.K.; Williams, B.A.; Dhanoa, M.S. et al. A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. Animal Feed Science and Technology, v. 48, p. 185-197, 1994.
Van Lier, E.; Regueiro, M. Digestión en retículo-rumen. Montevideo (Uruguay): Universidad de la República, 2008. Disponible en: http://prodanimal.fagro.edu.uy/cursos/AFA/ TEORICOS/Repartido-Digestion-en- Reticulo-Rumen.pdf. Accesado en: 25/04/2016.
Vargas, J.; Pabón, M.; Carulla, J. Producción de metano in vitro en mezcla de gramíneas-leguminosas del trópico alto colombiano. Revista Archivos de Zootecnia, v.63, n.243, p.397-407,2014.
Villalobos, L. Fenología, producción y valor nutritivo del pasto alpiste (Phalaris arundinacea) en la zona alta lechera de Costa Rica. Agronomía costarricense, v. 36, n.1, p.14, 2014.
Villalobos, L.; Sánchez, J.M. Evaluación agronómica y nutricional del pasto raygrass perenne tetraploide Lolium perenne producido en zonas altas de Costa Rica. Producción de biomasa y fenología. Agronomía Costarricense, v. 34, n.1, p. 31-42, 2010.
Como citar: Apráez-Guerrero, J.E.; Delgado-Jurado, D.; Solarte-Portilla, C. Producción de metano in vitro en las variedades de pasto ryegrass: sterling nui, ohau y samson. Revista Veterinaria y Zootecnia, v. 10, n. 2, p. 90-105, 2016. DOI: 10.17151/vetzo.2016.10.2.8
Este obra está bajo una Licencia de Creative Commons Reconocimiento CC BY
|







 | Today | 46 |
 | Yesterday | 574 |
 | This week | 3879 |
 | Last week | 4726 |
 | This month | 15393 |
 | Last month | 15809 |
 | All days | 1359460 |
Online Now: 9
Your IP: 18.117.8.177
MOZILLA 5.0,
Today: November 24, 2024
Your IP: 18.117.8.177
MOZILLA 5.0,
Today: November 24, 2024