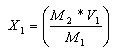|
Cinética de fermentación, pruebas de desafío in vitro y efecto de inhibición de Lactobacillus gasseri ATCC 199921 ARTÍCULO DE INVESTIGACIÓN
Henry Jurado-Gámez (Recibido: 10 de Junio de 2016 Aprobado: 18 de Noviembre de 2016 Actualizado: 13 de diciembre de 2016)
1 Financiado por la Vicerrectoría de Posgrados y Relaciones Internacionales (VIPRI), Universidad de Nariño, Pasto, Colombia. 2 Programa de Zootecnia, Departamento de Producción y Procesamiento Animal, Facultad de Ciencias Pecuarias, Universidad de Nariño, Pasto,Colombia. 3 Grupo de investigación en Procesos Biotecnológicos Aplicados a la Producción Animal - Fisiología y Etología Animal (PROBIOTEC-FISE), Pasto, Colombia.
This email address is being protected from spambots. You need JavaScript enabled to view it.
DOI: 10.17151/vetzo.2016.10.2.7
RESUMEN: Se evaluó el potencial in vitro de Lactobacillus gasseri, para ello, se determinó la susceptibilidad/resistencia a penicilina (P), cefalotina (KF), ciprofloxacina (CIP), gentamicina (CN) y dicloxacilina (DCX). Se estudió el efecto inhibitorio de L. gasseri y el sobrenadante sobre Y. pseudotuberculosis. El crecimiento de L. gasseri se estudió en diferentes condiciones de estrés como temperatura 38oC y 45oC; pH de 2,5-3,5-4,5 y 6; concentraciones de 3, 4 y 5% de sales biliares y 0,5-1 y 2% de bilis bovina. En la cinética se determinaron UFC/150µl, pH, acidez, consumo de azúcar y determinación de proteína en los medio MRS y PRO. Se realizó un análisis de péptidos y ácido láctico mediante cromatografía líquida de alta eficiencia. Se encontró que L. gasseri fue resistente a KF y DCX, a diferencia de Y. pseudotuberculosis que fue resistente únicamente a DCX. L. gasseri y su sobrenadante inhibió a la bacteria patógena, con halos de 4 y 3 ml. Se observaron crecimientos de 3x1013 y 3x1012 UFC/150µL a 38°C y 45°C; 3x109-1,3x1010 y 3x109 UFC/150µL a pH de 6-4,5 y 3,5; 2,7x1012-1,3x1012 y 2,5x1012 UFC/150µL a 3-4 y 5% de sales biliares; 5x1012-2,4x1012 y 3,5x1012 UFC/150µL en 0,5-1 y 2% de bilis bovina. La fase exponencial de los medios MRS y PRO ocurrió a las 20 y 16 horas con valores de 5x1011 y 3,7x1012 UFC/150µL; el pH registró valores iniciales y finales entre 5,72-4,6 y 5,7-4,48; porcentajes iniciales y finales de 0,54-1,32% y 0,21-0,90% de acidez. El consumo total de azúcares en los medios MRS y PRO fue de 69,94 y 88,09% y 0,66 y 3,12 mg/L de proteína. El sobrenadante registró valores de 4,80 y 5,10 g/L de ácido láctico y un péptido con una concentración de 0,58 mg/ml.
Palabras clave: viabilidad, resistencia, probiótico, patógeno, fermentación
Fermentation kinetics, in vitro challenge tests and inhibitory effect of Lactobacillus gasseri ATCC 19992
ABSTRACT: The potential in vitro of Lactobacillus gasseri was evaluated. To do that, the susceptibility/resistance to penicillin (P), cephalothin (KF), ciprofloxacin (CIP), gentamicin (CN) and dicloxacillin (DCX) was determined. The inhibitory effect of L. gasseri and the supernatant on Y. pseudotuberculosis was studied. L. gasseri growth was studied in various stress conditions such as temperature 38oC and 45oC, pH 2.5-3.5-4.5 and 6, concentrations of 3-4 and 5% bile salts and 0.5-1 and 2% of bovine bile. In the kinetics CFU/150μl, pH, acidity, sugar consumption and protein determination on MRS and PRO mediums were determined. An analysis of peptide and lactic acid was performed using high-performance liquid chromatography. It was found that: L. gasseri was resistant to KF and DCX unlike Y. pseudotuberculosis was resistant only to DCX. L. gasseri and its supernatant inhibited the pathogenic bacteria with halos of 4 and 3 ml; growths of 3x1013 and 3x1012 CFU/150μl at 38°C and 45°C; 3x109-1,3x1010 and 3x109 CFU/150μl at pH from 6-4.5 and 3.5; 2.7x1012-1.3x1012 and 2.5x1012 CFU/150μl to 3-4 and 5% bile salts; 5x1012-2.4x1012 and 3.5x1012 CFU/150μl to 0.5-1 and 2% bovine bile were observed. The exponential phase of the MRS and PRO mediums occurred at 20 and 16 hours with values of 5x1011 and 3.7x1012 CFU/150μl; the pH recorded initial and final values between 5.72 to 4.6 and from 5.7 to 4.48, and initial and final rates of 0.54 to 1.32% and 0.21 to 0.90% acidity. The total consumption of sugars in the MRS and PRO mediums was 69.94 and 88.09% and 0.66 and 3.12 mg/L of protein. The supernatant recorded values of 4.80 and 5.10 g/L of lactic acid and a peptide with a concentration of 0.58 mg/ml.
Key words: viability, resistance, probiotic, pathogen, fermentation
Introducción
Los probióticos son principalmente bacterias no patógenas, que pueden ser utilizados como suplemento alimenticio, los cuales tras ser ingeridos en cantidades adecuadas, pueden mejorar el equilibrio microbiano y producir efectos benéficos en la salud de quienes los ingieren (Manzano et al., 2012).
Las bacterias del género Lactobacillus son Gram positivas, no generan esporas y algunas cepas presentan cuerpos bipolares (Fuenmayor, 2009). Este género muestra una gran diversidad de especies difundidas en la naturaleza, lo cual se debe a su capacidad de crecer en diferentes condiciones medioambientales (Ramírez-Ramírez et al., 2011).
Las condiciones de crecimiento del género Lactobacillus son variadas; cada especie de este género presenta particularidades respecto a los requerimientos de aminoácidos, péptidos, derivados de ácidos nucleicos, vitaminas, sales, ácidos grasos o ésteres de ácidos grasos y carbohidratos (Jiménez, 2010).
“Lactobacillus gasseri es una especie caracterizada de bacterias Gram positivas, bajas en guanina y citosina (GC), conocidas por representar uno de los principales Lactobacillus homofermentativo del intestino humano” (Fakhry apud Di Luccia et al., 2013, p 8). L. gasseri se encuentra asociada a diversas funciones probióticas que incluyen la disminución de la actividad de las enzimas fecales mutagénicas, estimulación de los macrófagos, adhesión a los tejidos intestinales, y la producción de bacteriocinas capaces de disminuir organismos patógenos (Anwar et al., 2010).
Y. pseudotuberculosis causa una zoonosis bacteriana denominada yersiniosis y puede causar infecciones zoonóticas animales salvajes, domésticos y aves (Kasper & Faud apud Amaya & Calle, 2008; Galindo et al., 2011).
La investigación tuvo como objetivos establecer el efecto de inhibición de L. gasseri y su sobrenadante sobre Y. pseudotuberculosis, estudiar el crecimiento de L. gasseri bajo diferentes condiciones de estrés (temperatura, pH, sales biliares y bilis bovina) e identificar las condiciones de la cinética de fermentación de L. gasseri en el medio MRS y PRO.
Materiales y Métodos
Esta investigación se realizó en los laboratorios de la Facultad de Ciencias Pecuarias y los laboratorios especializados pertenecientes a la Universidad de Nariño, ubicados en la ciudad de Pasto con una temperatura promedio de 14°C, a una altura de 2540 msnm, precipitación anual promedio de 1084 mm y humedad relativa del 76%.
Se utilizó la cepa de colección American Type Culture Collection (ATCC) Lactobacillus gasseri ATCC® 19992™*- KS2C y la cepa de colección National Collection of Type Cultures (NCTC) Yersinia pseudotuberculosis NCTC 8580, ambas fueron reconstituidas de acuerdo con las indicaciones de la casa comercial. L. gasseri se conservó mediante repique en medio sólido MRS (Man, Rogosa y Sharpe) cada 5 días y en medio líquido MRS cada 8 días. En el caso de Y. pseudotuberculosis fue conservada en caldo BHI (Brain-heart infusión) y agar MacConkey como medio sólido, las condiciones de incubación fueron a 37°C y 24 horas, para luego ser refrigeradas (4°C) hasta su utilización.
Para cultivar el inóculo se tomó una alícuota de la cepa láctica y se sembró con anterioridad en cajas de agar MRS con azul de anilina y se transfirió a un Erlenmeyer (100 ml), que contenía 40 ml de caldo MRS estéril; posteriormente fue llevado a incubación durante 24 horas a una temperatura de 37°C. Trascurridas las 24 horas, con una pipeta de 5 ml, se transfirieron 4 ml del contenido del Erlenmeyer y se trasladó a otro Erlenmeyer con 40 ml de caldo MRS y se incubó por 24 horas a 37°C.
Se ajustó el inóculo a 10% v/v para iniciar la fermentación. Luego de las 24 h se calculó el número de bacterias por ml. El patrón en la escala de McFarland a la cual se ajustó la población bacteriana fue de 4 (1,2 x 109 UFC/ml). Con la ayuda de una pipeta, se tomó 1 ml del contenido del Erlenmeyer y se procedió a hacer la lectura en espectrofotómetro a 625 nm (Crueger & Crueger, 1993). En los casos en que se presentó mayor población de la establecida, se adicionó caldo MRS estéril teniendo en cuenta el siguiente cálculo matemático de proporcionalidad de acuerdo con Guerrero apud Montes et al. (2003).
M1= Población o densidad celular que se desea ajustar. M2= Densidad óptica utilizada en la primera fermentación. V1= 1 ml Volumen proveniente del inóculo total (10/90). X1= Cantidad que contiene M2 V2= Volumen que se agrega a un (1) ml para ajustar a 1,50 x 108 UFC/ml. V3= 100 ml Cantidad total del inóculo. X2= Cantidad de caldo MRS comercial estéril que se agrega a V3 para ajustar la población al valor de M2. Encontramos entonces X1. M1-----M2 M2-----X1
Se calcula V2; V2=V1-X1 Se determina el valor de X2 V1------V2 V3------X2
X2, corresponde al volumen que debe añadirse a 100 ml para ajustar la población.
Se realizaron pruebas de susceptibilidad a antibióticos en L. gasseri y Y. pseudotuberculosis; los antibióticos evaluados fueron Gentamicina (CN 10 µg), Penicilina (P10 IU), Ciprofloxacina (CIP 5 µg), Dicloxacilina (DCX 1 µg) y Cefalotina (KF 30 µg). Se utilizó el método de Kirby & Bauer (1966); en donde las bacterias fueron depositadas en un 1 ml de agua destilada, por separado y fueron incubados a 37°C, hasta lograr una turbidez similar al estándar 0,5 de MacFarland; posteriormente, se transfirió a cajas de petri con agar Mueller Hilton y se difundió con un hisopo de algodón. Luego, los discos de los antibióticos se colocaron sobre el agar, haciendo presión para fijarlos al medio; posteriormente se invirtieron las cajas de petri y se incubaron a 37°C por 18 h; al término de las 18 h, se midieron los halos formados entre el borde del disco y el borde máximo de inhibición.
De igual manera, se evaluó el efecto de inhibición producido por L. gasseri sobre Y. pseudotuberculosis mediante la metodología de Tagg & McGiven (1971). Se ajustó una alícuota de L. gasseri a escala McFarland 0,5 y se cultivaron en agar MRS más azul de anilina a concentraciones de 50, 75, 100 y 150µl; finalmente se incubaron a 37°C por 24 h. La bacteria patógena se ajustó a escala MacFarland 0,5 (1,5 x 108 UFC/ml) y se cultivó en agar TSA (Triptona soya agar). Se hicieron discos a partir del agar MRS más azul de anilina, en donde se sembró L. gasseri y fueron puestos en las cajas con la bacteria patógena, luego se incubó a 32°C por 12 horas. Al finalizar el periodo de incubación se midieron los halos de inhibición. Se determinó susceptibilidad de la cepa patógena cuando el halo fue igual o superior a 2 mm (Ramírez, 2005).
El sobrenadante de L. gasseri se ajustó a 4 en la escala McFarland (625 nm). Posteriormente se depositaron muestras de 1,5 ml en tubos Eppendorf y se centrifugaron a una temperatura de 4°C con 15000 rpm durante 15 min. Se utilizó el sobrenadante filtrándolo con papel filtro de 0,25 µm y sin filtrar. Cada muestra se refrigeró a 4°C para posterior análisis (Jurado-Gámez, 2014a).
Los sobrenadantes se evaluaron mediante el método de Kirby & Bauer (1966) modificado, con dos métodos diferentes: en el primero se adicionó el sobrenadante a discos de celulosa, en concentraciones de 50, 75, 100 y 150 µl, y en el segundo se utilizaron cilindros estériles de 6 mm de diámetro, en donde se depositaron las mismas concentraciones. Los discos y cilindros fueron colocados en cajas de Petri con la cepa patógena y se incubaron a 37°C por 24 h.
L. gasseri se estudió a concentraciones de 3, 4 y 5% de sales biliares bovina y concentraciones de 0,5 - 1 y 2% de bilis bovina. Se partió del cultivo del inoculo; de este cultivo se tomaron alícuotas que fueron sembradas en caldo MRS con las diferentes concentraciones de sales biliares y bilis bovina de manera separada, luego se tomaron muestras y se cultivaron en agar MRS con azul de anilina, permaneciendo 48 horas a 32°C.
Se realizaron pruebas de producción de gas y catalasa a la bacteria láctica (Cai et al. 1999) y el crecimiento de L. gasseri se evaluó a pH 2,5- 3,5- 4,5 y 6, este ensayo se realizó durante tres horas, tomando muestras cada 20 minutos. La cepa fue incubada en medio MRS comercial y el pH se ajustó con la adición de ácido tartárico, para inhibir el efecto de la producción de ácido láctico. Las condiciones de incubación fueron de 32°C por 48 h.
La cinética de fermentación de L. gasseri se evaluó en los medios de cultivo MRS y PRO (compuesto por 10 g/l azúcar blanco, 15 g/l leche de soya, 150 g/l suero de leche, 15 g/l salvado de trigo, Ramírez 2005), cada medio se dispuso en un Erlenmeyer con 540 ml de medio y 60 ml de inóculo y se incubó en Shaker en agitación constante a 37°C y 100 rpm. Durante la cinética se evaluó: conteo de microorganismos viables en placa (UFC/ml), pH, consumo de azúcar total y proteína, determinación de acidez y determinación de producción de ácido láctico. Las mediciones se realizaron cada 4 h durante 24 h.
Para el conteo de microorganismos viables en placa, se diluyó 1 ml de muestra en 9 ml de agua peptonada al 0,1%, se realizaron diluciones decimales que fueron transferidas a cajas de Petri, que contenían medio MRS con azul de anilina (0,1 ml) para siembra en superficie. Las cajas fueron incubadas a 37°C y se observaron entre 24 y 48 horas. Se tomaron como criterio de inclusión las cajas con conteos entre 30 y 300 colonias. El número de colonias fue multiplicado por el inverso de la dilución y por 10 para obtener UFC/150µl (Jurado-Gámez et al., 2014b). Para determinar el pH se tomó una muestra del medio y se midió con pHmetro digital (JENCO® VisionPlus).
El método de Dubois et al. (1956) fue usado para determinar el azúcar total, se prepararon diferentes concentraciones de glucosa para crear una curva patrón mediante los valores obtenidos de las muestras observadas a una densidad óptica de 625 nm. Los valores se graficaron contra la concentración en mg/l, finalmente se obtuvieron los valores de la línea recta.
El ácido láctico fue determinado mediante titulación con hidróxido de sodio (1N). La biomasa se determinó por los métodos de Crueger & Crueger (1993) y Rodríguez et al. (2003); para ello se estableció la velocidad máxima de crecimiento mediante la siguiente ecuación:
El tiempo de duplicación celular (td) se determinó teniendo en cuenta la siguiente ecuación:
Se cuantificó el consumo de proteína por el método de Lowry et al. (1951) con modificación de Malara & Charra (1972), se realizó una curva de calibración mediante seroalbúmina bovina y se determinó la absorbancia en espectrofotómetro a 625 nm. Los valores obtenidos fueron graficados contra la concentración para obtener la ecuación de la línea recta.
Se evaluó la viabilidad de L. gasseri a dos temperaturas 38 y 45°C, tomando como referencia la fase exponencial de crecimiento encontrada en la cinética de fermentación, el procedimiento se basó en lo propuesto por Crueger & Crueger (1993), para ello se ajustó el inóculo de acuerdo con la escala de MacFarland 0,125; la incubación duró 12 h de iniciada la prueba, enseguida se realizaron diluciones de 10-1 hasta 10-12 en agua peptonada y se sembraron en cajas de Petri con azul de anilina comenzando en la disolución de 10-6 hasta la máxima disolución; las cajas se incubaron por 48 horas a 37°C para determinar el recuento de colonias en UFC/150 µl.
Se identificó el contenido de péptidos en el sobrenadante mediante HPLC. Se tomó 1 ml de sobrenadante y se filtró con jeringa PVDF, Pall de 0,25 µm. La muestra se congeló (-20°C) y protegió de la luz hasta su análisis. Se identificaron los espectros UV de los picos integrados, mediante el sistema de detección PDA y se compararon con el patrón de péptidos Sigma H2016, se realizaron 4 réplicas.
El diseño estadístico utilizado fue el de medidas repetidas en el tiempo, y se usó el paquete estadístico SAS 9.1 (2004), con el procedimiento PROC MIXED de SAS.
Resultados y Discusión
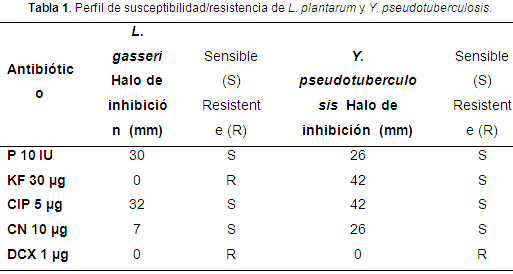
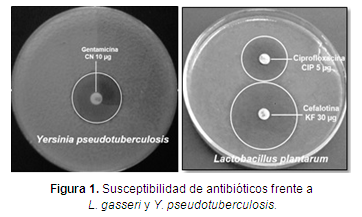
Los resultados del perfil de susceptibilidad/resistencia de L. gasseri mostró resistencia a la acción de KF y DCX y sensibilidad a la P, CIP y CN (tabla 1, figura 1).
Se debe procurar que las bacterias benéficas no presenten resistencia a los antibióticos, para evitarse el riesgo de transmisión de genes de resistencia a bacterias patógenas que estén presentes en el TGI del huésped (Sanz et al. apud Alvarado & Díaz, 2009).
Según Morillo & Romero (2016), L. gasseri presentó resistencia a KF y DCX, lo cual concuerda con los resultados reportados en esta investigación. Por su parte, Rubio et al. (2013) reporta la resistencia de L. gasseri a kanamicina y clindamicina. El análisis de la secuencia genómica de L. gasseri ATCC 33323 permitió inferir que no posee genes de resistencia a antibióticos que puedan ser transmisibles, pero hace la aclaración, de que no es correcto suponer que todas las cepas de L. gasseri estén libres de genes de resistencia a antimicrobianos transmisibles (Azcárate et al. apud Selle & Klaenhammer, 2008).
Y. pseudotuberculosis mostró resistencia a DCX (tabla 1, figura 1), resultados similares fueron reportados por Jurado et al. (2014a) y Morillo & Romero (2016).
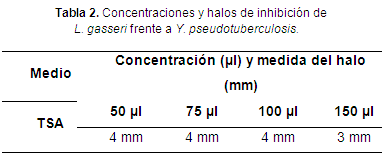
L. gasseri inhibió a Y. pseudotuberculosis con halos de 3 y 4 mm (tabla 2 y figura 2).
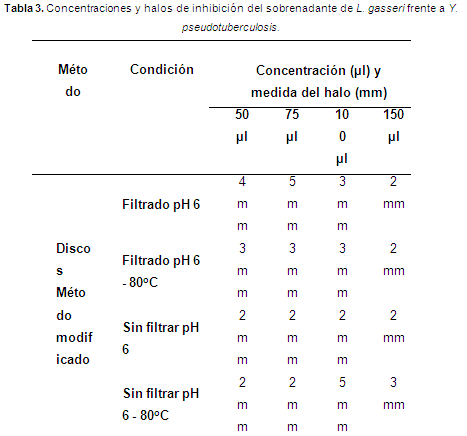
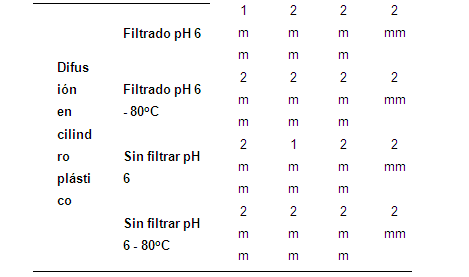
Las BAL pueden producir diferentes compuestos con efectos antimicrobianos (Mishra & Lambert apud Delgado (2005). Esta característica se utiliza para la destrucción de bacterias indeseables o patógenas y por lo general estos compuestos son ácido láctico o derivados del metabolismo del oxígeno como el H2O2 (Leveau & Bouix apud Peñaflor, 2007). Con base en los resultados registrados en la tabla 3, se infiere que L. gasseri inhibió a Y. pseudotuberculosis por acción individual o conjunta de los metabolitos producidos por L. gasseri.
Se observaron halos de inhibición generados por el sobrenadante entre 1 y 5 mm (Tabla 3).
Ya que las BAL generan diversos compuestos con actividad antimicrobiana, se dificulta estimar cuál de ellos tiene mayor o menor incidencia sobre el efecto de inhibición, por tal razón se eliminó del sobrenadante los posibles efectos de inhibición generados por compuestos como ácido láctico y biomasa bacteriana. Por lo tanto, de la información reportada en la tabla 4, se infiere que el sobrenadante inhibió a Y. pseudotuberculosis como consecuencia de las bacteriocinas en el sobrenadante.
Resistencia bajo condiciones de estrés (temperatura, sales biliares, bilis y pH): el ensayo de crecimiento a diferentes temperaturas reportó crecimientos máximos de 3x1013 y 3x1012 UFC/150µl a 38oC y 45oC respectivamente; por su parte, Gregoret et al. (2012) mencionó que una cepa de L. gasseri presentó un crecimiento de 108 UFC/ml a 37°C. Según Ahmed et al. (2006), las BAL tienen una temperatura de crecimiento óptimo y es una de las características que permite diferenciarlas, debido a que inciden en el crecimiento y puede afectar el tiempo de generación de acuerdo con la fase de crecimiento y especie. Es posible inferir que la temperatura óptima para L. gasseri se encuentra cercana a 38oC.
En cuanto al crecimiento en condiciones variables de pH, se registraron valores de 3,8x109, 3x1011 y 3x108 UFC/150 µl a pH de 6-4,5 y 3,5 después de 100 minutos; finalmente se encontraron 3x109-1,3x1010 y 3x109 UFC/150 µl a pH de 6-4,5 y 3,5 respectivamente en el último tiempo del ensayo (180 minutos). Por lo anterior se infiere que L. gasseri puede soportar una simulación de ambientes ácidos in vitro, manteniendo conteos viables de crecimiento durante 3 horas.
Por último, los resultados para sales biliares estuvieron comprendidos entre 4x108 y 2,7x1012 - 3,5x108 y 1,3x1012 y 2x107 a 2,5x1012 UFC/150 µl al 3, 4 y 5%. Po su parte, los valores de bilis bovina fueron de 3x108 y 5x1012 - 3,5x107 y 2,4x1012 y 4x108 a 3,5x1012 UFC/150 µl en 0,5-1 y 2% de bilis bovina. Gibson & Fuller apud Moreno, (2012) mencionan que las bacterias probióticas, antes de llegar al TGI, deben resistir inicialmente la acidez gástrica y posteriormente se deben enfrentar a las sales biliares. Con base en lo anterior y los resultados encontrados, L. gasseri tendría la capacidad de mantener conteos viables en condiciones simuladas de acidez, seguidas de porcentajes variables de sales biliares.
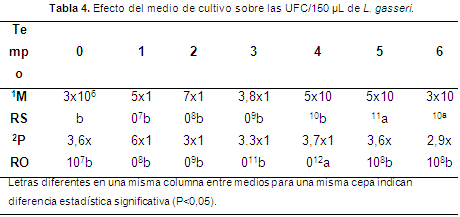
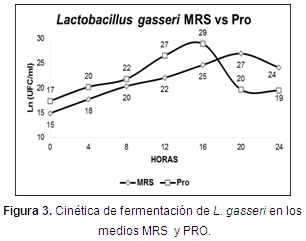
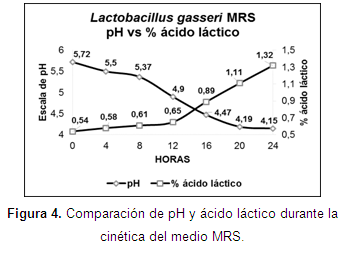
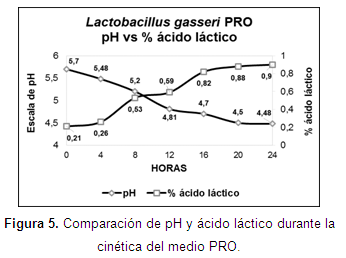
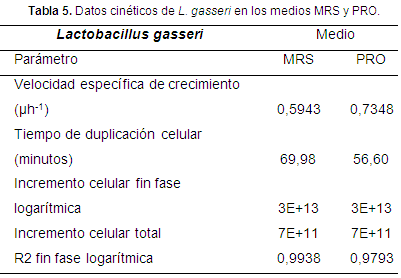
Cinética de fermentación: la fase exponencial en los medios MRS y PRO se encontró en los tiempos 5 (20 h) y 4 (16 h) (Figura 3), con valores de 5x1011 y 3,7x1012 UFC/150 µl. Se encontraron diferencias estadísticas significativas (P<0,05) desde el tiempo 3 hasta el tiempo 6 (Tabla 4).
Según Morillo & Romero (2016), L. gasseri reportó un valor de 1,75x1012 UFC/150 µl en el medio MRS y 6,2x1012 UFC/g en el medio MRS según Yoda et al. (2014). Estos resultados, comparados con los encontrados en esta investigación, muestran que los dos medios tuvieron crecimientos adecuados, sin embargo la fase exponencial del medio PRO fue más rápida. Según Orozco & Solarte apud Morillo & Romero (2016), en la fermentación las BAL tienen que suplir sus requerimientos nutricionales (carbohidratos, proteínas, lípidos y vitaminas), ya sea del medio de cultivo o por la interacción con los microorganismos presentes en el medio de fermentación. Según esto, el medio PRO fue capaz de suplir de forma eficiente los requerimientos de L. gasseri en comparación con el medio MRS.
Los valores de pH y ácido láctico, mostraron una relación inversamente proporcional en los dos medios (Figuras 4 y 5). El efecto de la biomasa (UFC/150µl) sobre el pH del medio de cultivo se presentaron diferencias estadísticas significativas (P<0,05), destacándose un menor pH para el medio MRS (4,23 y 4,20) vs PRO (4,53 y 4,50). En cuanto al efecto de biomasa sobre el porcentaje de ácido láctico del medio de cultivo, se observaron diferencias estadísticas significativas (P<0,05) en los tiempos 1, 2, 4, 6 y 7 con mayor porcentaje de ácido en el medio MRS (0,60; 0,64; 0,73; 1,21; 1,46) vs PRO (0,23; 0,28; 0,60; 0,97; 0,97).
Ahmed et al. (2006) afirman que existe una relación inversa entre la acidez y el pH; por su parte, Doelle (2014) menciona que los nutrientes son absorbidos mientras avanza el crecimiento celular hacia la fase de crecimiento logarítmico y los productos finales se excretan, haciendo que esta actividad metabólica altere el pH y cambie el entorno. Esta relación se observó en los dos medios de cultivo evaluados, por lo cual se infiere que los nutrientes fueron consumidos provocando una excreción principalmente de ácido láctico, que cambió el pH de los medios.
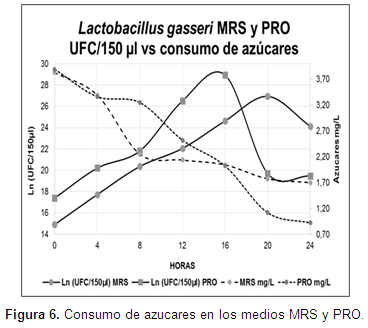
El consumo de azúcar presentó valores iniciales y finales de 3,83 y 1,70 mg/l en el medio MRS y 3,89 y 0,92 en el medio PRO.
Los valores iniciales y finales demuestran que L. gasseri mantuvo un consumo de azúcares progresivo (Figura 6). Al respecto, Cardelle et al. (2011) mencionan que en la fermentación in vitro de cepas de lactobacilos utilizadas en combinación con diferentes materias primas como sustrato (prebióticos), se dificulta identificar cuáles son fermentados selectivamente por los microorganismos de interés, y también se dificulta establecer la velocidad a la que un solo oligosacárido es fermentado. Lo anterior, se encuentra en concordancia con los resultados encontrados, ya que es posible identificar el consumo progresivo de azúcar, pero se dificulta establecer el tipo de carbohidrato que L. gasseri consumió selectivamente. En contraste, es importante identificar aquellos carbohidratos que las BAL son incapaces de fermentar; según Anwar et al. (2010), tres cepas de L. gasseri, reportaron la imposibilidad de fermentar rafinosa.
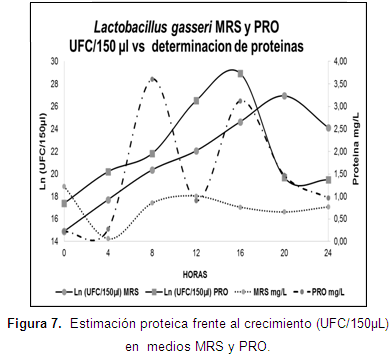
La determinación de proteínas en la fase exponencial para L. gasseri en los medios MRS y PRO fue de 0,66 y 3,12 mg/l. Esta determinación no presenta curvas con tendencia definida, a diferencia de la curva correspondiente al consumo de azúcares (Figura 7).
Elías et al, apud Chafla et al., (2015) mencionan que el incremento de la concentración proteica en un medio, se encuentra profundamente asociado al desarrollo de bacterias y enzimas. Por tal razón, las variaciones de proteína registradas en la gráfica 7, posiblemente indiquen un consumo de proteína del medio, la producción de enzimas y la estimación indirecta de la biomasa bacteriana.
El medio PRO en comparación con el medio MRS, reportó ser más eficiente en cuanto a velocidad específica de crecimiento (µh-1) y tiempo de duplicación celular.
La velocidad de crecimiento específica puede alcanzar valores máximos cuando la concentración de nutrientes es alta (Orozco & Solarte apud Morillo & Romero, 2016). Los resultados encontrados en cuanto a µh-1 y tiempo de duplicación celular, permiten inferir que posiblemente el medio PRO fue el medio que suplió los requerimientos nutricionales de L. gasseri de forma eficiente en cuanto a tipo de requerimiento y cantidad; resultados similares fueron descritos por Calpa & Chaspuengal (2013), referentes al menor tiempo de duplicación y un mayor µh-1 para Lactobacillus casei en un medio similar al medio PRO.
Análisis de HPLC en muestras de sobrenadante: El análisis del sobrenadante de L. gasseri registró una cantidad de ácido láctico de 4,80 y 5,10 g/l y un péptido conformado por una cadena de aminoácidos VAL-TIR-VAL, con una concentración de 0,58 mg/ml.
Ramírez (2008) menciona que entre los principales metabolitos producidos por las BAL, se encuentran los compuestos de bajo peso molecular y sustancias con efecto inhibitorio sobre otros microorganismos; el principal de estos compuestos es el ácido láctico, producido por la fermentación de las bacterias, el cual puede reducir el pH hasta un pH que otras bacterias no toleran.
Fujimura et al. (2012) encontraron que L. gasseri OLL 2716 produjo 1,4 g/l de ácido láctico, cantidad que fue menor a la registrada en este estudio. Esta variación puede estar influenciada por diferencias entre la cepa bacteriana, temperatura y el medio de crecimiento. Waldir et al. (2007) mencionan que la concentración de la fuente de carbono marca significativamente en la conversión de glucosa en ácido láctico.
El análisis de péptidos detectó 7 picos en el sobrenadante de L. gasseri, los cuales fueron comparados con un patrón estándar, el cual reportó una similitud entre el pico 6 equivalente al tiempo de retención 11,95; por la cual se estableció que posiblemente se trató de un péptido conformado por una cadena de aminoácidos VAL-TIR-VAL.
Conclusiones
La simulación in vitro de L. gasseri permitió establecer que podría resistir el paso a través del TGI y soportar las condiciones fisiológicas de temperatura, acidez gástrica y enzimas en cantidades viables. Además, cuenta con mecanismos para inhibir a bacterias patógenas y potencialmente se evita el riesgo de transmitir genes de resistencia a P, CIP y CN.
Referencias bibliográficas
Alvarado-Rivas, C. y Díaz-Rivero, C.G. Estudio preliminar del potencial probiótico lactobacilos aislados de pastizal de una finca lechera. Revista de la Facultad de Farmacia, v. 51, no.1, p. 8-14, 2009.
Amaya-Garzón, M.A.; Calle-Londoño, L. Avances en la producción de una vacuna viva contra Yersinia pseudotuberculosis y evaluación de su efectividad mediante un ensayo de infección experimental en Cavia porcellus. Bogotá, Colombia: Universidad de La Salle, 2008. 49p. Tesis (Medicina Veterinaria).
Anwar, M.A.; Kralj, S.; Piqué, A.V.; Leemhuis, H et al. Inulin and levan synthesis by probiotic Lactobacillus gasseri strains: characterization of three novel fructan sucrase enzymes and their fructan products. Microbiology, v. 156, n. 4, p. 1264-1274, 2010.
Bauer, A.; Kirby, J.; Sherris, T. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol., v. 45, n. 4, p. 493-496,1966.
Cai, Y.; Suyanandana, P.; Saman, P.; Benno, Y. Classification and characterization of lactic acid bacteria isolated from the intestines of common carp and freshwater prawns. J Gen Appl Microbiol., v. 45, n. 4, p. 177-184, 1999.
Calpa-Yamá, F.Y.; Chaspuengal-Tulcán, A.M. Evaluación in vitro de Lactobacillus casei con características probióticas sobre Yersinia pseudotuberculosis. San Juan de Pasto, Colombia: Universidad de Nariño. 2013, 107p. Tesis (Zootecnia).
Cardelle, A.; Corzo, N.; Olano, A. et al. Galactooligosaccharides derived from lactose and lactulose: influence of structure on Lactobacillus, Streptococcus and Bifidobacterium growth. International Journal of Food Microbiology, v. 149, p.1 2-34, 2011.
Chafla, A.L.; Espin, J.M.; Jara, M. et al. Caracterización de proteína microbiana obtenida a partir de residuos agroindustriales. Revista Amazónica Ciencia y Tecnología, v. 4, n. 2, p. 131-145, 2015.
Crueger, W.; Crueger, A. Biotecnología: Manual de Microbiología Industrial. 3 ed. España: Ed. Acribia, 1993, 413p.
Delgado-Palacio, S. Microbiota intestinal humana: análisis y evolución de poblaciones representativas e identificación de bacterias probióticas. Oviedo: Instituto de Productos Lácteos de Asturias, 2005. 162p. (Memoria para optar al Grado de Doctor).
Di Luccia, B.; Manzo, N.; Baccigalupi, L. et al. Lactobacillus gasseri SF1183 Affects Intestinal Epithelial Cell Survival and Growth. Journal. pone., v.8, n.7, p. 8, 2013.
Doelle, H.W. Bacterial metabolism. Academic Press, 2. ed. Australia: St. Lucia, Brisbane, Academic Press, INC. 2014, 738p.
Dubois, M.; Gilles, K.: Hamilton, J. et al. Colorimetric method for determination of sugar and related substances. Anal Chem., v.28, p. 350-356, 1956.
Fuenmayor-Bobadilla, C.A. Aplicación de bioprocesos en polen de abejas para el desarrollo de un suplemento nutricional proteico. Medellín, Colombia: Universidad Nacional de Colombia. 2009. 134p. Tesis (Maestría en Ingeniería Química Área de Bioprocesos)
Fujimura, S.; Watanabe, A.; Kimura, K.; Kaji, M. Probiotic mechanism of Lactobacillus gasseri OLL2716 strain against Helicobacter pylori. Journal of clinical microbiology, v.50, n.3, p. 1134-1136, 2012.
Galindo, C.L.; Rosenzweig, J.A.; Kirtley, M.L. et al. Pathogenesis of Y. enterocolitica and Y. pseudotuberculosis in Human Yersiniosis. Journal of pathogens, p.16, 2011.
Gregoret, V.; Perezlindo, M.; Vinderola, G. et al. A comprehensive approach to determine the probiotic potential of human-derived Lactobacillus for industrial use. Food Microbiology, v.34, p. 19-28. 2012.
Jiménez-Serna, A. Efecto de diferentes fármacos sobre el crecimiento de Lactobacillus casei shirota. México D.F. Instituto politécnico nacional de ciencias biológicas. 2010, p.12-13. Tesis (Tesis Doctoral)
Jurado-Gámez, H.; Calpa-Yama, F.; Chaspuengal-Tulcán, A. Determinación in vitro de la acción probiótica de Lactobacillus plantarum sobre Yersinia pseudotuberculosis aislada de Cavia porcellus. Revista Facultad Medicina Veterinaria y Zootecnia, v.61, n.3 p. 241-257, 2014a.
Jurado-Gámez, H.; Martínez, J.A.; Chaspuengal, A.M. et al. Evaluación in vitro de la acción de Lactobacillus plantarum con características probióticas sobre Yersinia pseudotuberculosis. Biotecnología en el Sector Agropecuario y Agroindustrial, v.12, n. 2, p. 49-59, 2014b.
Manzano, C.; Estupiñán, D.; Poveda, E. Efectos clínicos de los probióticos: qué dice la evidencia. Revista chilena de nutrición, v.39, n.1, p. 98-110, 2012.
Malara, G.; Charra, R. Dosage des proteines particulaires selon la methode the Lowry. Notes de Travail. 5: 11. Station Zoologique de Vellafranche-Sur-Mer, France Universidad de París. 1972
Montes, A.; Santacruz, A.; Sañudo, J. Efecto in vitro de Lactobacillus casei subsp. rhamnosus sobre el crecimiento de un aislado de Helicobacter pylori. San Juan de Pasto, Colombia: Universidad de Nariño. 2003, 120p. Tesis (Microbiología)
Morillo-Garcés, J.A.; Romero-Benavides, D.A. Evaluación in vitro del efecto probiótico de Lactobacillus gasseri ATCC 19992 y lactococcus lactis subsp. lactis ATCC 11454 sobre Yersinia pseudotuberculosis NCTC 8580. San Juan de Pasto, Colombia: Universidad de Nariño. 2016, 133p. (Tesis Zootecnia).
Moreno-Galarza, L.J. Aislamiento y Selección de Lactobacillus sp con potencial probiótico a partir de pan de abejas. Bogotá, Colombia: Universidad Nacional de Colombia, 2012. 109p. (Magíster en Ciencias-Microbiología).
Lowry, N.J.; Rosebrough, A.L.; Farr, L.; Randall. R.J. Proteine asurement with the folinphenol reagent.J. Biol. Chem. 1951, p.265-275. 1966.
Peñaflor-Mora, N.; García-Guerrero, A. Susceptibilidad de bacterias ácido lácticas (BAL) frente a diversos antibióticos. Pachuca de Soto, Hidalgo. Universidad Autónoma del estado de Hidalgo, 2007. 113p. Tesis (Medicina veterinaria).
Ramírez-Chavarín, N.L.; Wacher-Rodarte, C; Pérez-Chabela, M.L. Characterization and identification of thermotolerant lactic acid bacteria isolated from cooked sausages as bioprotective cultures. Journal of Muscle Foods, v.21, p. 585–596. 2008.
Ramírez-Cuenca, M. Actividad inhibitoria de cepas de bacterias ácido lácticas frente a bacterias patógenas y deterioradoras de alimentos. Hidalgo, México: Universidad Autónoma del Estado de Hidalgo. 2005. 77p. Tesis (Microbiología).
Ramírez-Ramírez, J.; Rosas-Ulloa, P.; Velásquez-González, M. Bacterias lácticas: importancia en alimentos y sus efectos en la salud. Revista Fuente, v. 2 p. 1-16, 2011.
Rodríguez, l.; Bueno, G.; Rodríguez, D. et al. True and apparent yields and maintenance coefficient and their significance on fermentation kinetics. New Horizons Biotechnology, p.163-172, 2003.
Selle, K.; Klaenhammer. T. Genomic and phenotypic evidence for probiotic influences of Lactobacillus gasseri on human health. FEMS microbiology reviews, v. 37, p. 915-935, 2013.
Tagg J.; McGiven, A. Assay system for bacteriocins. Appl. Environ. Microb., v. 2, p. 282-283, 1970.
Waldir, E.; Rychtera, M.; Melzoch, K. et al. Producción de ácido láctico por Lactobacillus plantarum L10 en cultivos batch y continuo. Revista Peruana de Biología, v. 14, no. 2, p. 271-276. 2007.
Yoda, K.; He, F.; Kawase, M. et al. Oral administration of Lactobacillus gasseri TMC0356 stimulates peritoneal macrophages and attenuates general symptoms caused by enteropathogenic Escherichia coli infection. Journal of Microbiology, Immunology and Infection, v.47, p. 81-86, 2014.
Como citar: Jurado-Gámez, H.; Martínez-Benavides, J.; Romero-Benavides, D.A.; Morillo-Garcés, J.A.; Orbes-Villacorte, A.E.; Mesías-Pantoja, L.N. Cinética de fermentación, pruebas de desafío in vitro y efecto de inhibición de Lactobacillus gasseri ATCC 19992. Revista Veterinaria y Zootecnia, v. 10, n. 2, p. 72-89, 2016. DOI: 10.17151/vetzo.2016.10.2.7
Este obra está bajo una Licencia de Creative Commons Reconocimiento CC BY
|





 | Today | 105 |
 | Yesterday | 574 |
 | This week | 3938 |
 | Last week | 4726 |
 | This month | 15452 |
 | Last month | 15809 |
 | All days | 1359519 |
Online Now: 11
Your IP: 3.145.91.111
MOZILLA 5.0,
Today: November 24, 2024
Your IP: 3.145.91.111
MOZILLA 5.0,
Today: November 24, 2024