|
Estructura genética de una población de bovinos Holstein en el departamento de Antioquia, usando polimorfismos del gen del factor de crecimiento insulínico tipo 2 (IGF2a)1
ARTÍCULO DE INVESTIGACIÓN
Melissa García-Valencia
1Grupo BIOGEM, Universidad Nacional de Colombia sede Medellín. 2Profesor Universidad Nacional de Colombia, sede Medellín, Facultad de Ciencias Agrarias, Departamento de Producción Animal, Grupo BIOGEM.
This email address is being protected from spambots. You need JavaScript enabled to view it.
Recibido: 23 de Agosto de 2016; aprobado 21 de Abril de 2017, Actualizado: 13 de Junio de 2017
DOI: 10.17151/vetzo.2017.11.1.2
RESUMEN
Desde los inicios de la tecnificación del campo, el objetivo de la producción pecuaria ha sido el perfeccionamiento de los sistemas por medio de la optimización de los recursos para alcanzar alto nivel productivo. No todos los componentes biológicos de dichos sistemas responden de igual manera a los nutrientes ofrecidos, esto en gran parte debido a su componente genético. Como respuesta a esta situación ha surgido una herramienta de gran utilidad que se podría implementar y aplicar al manejo diario de los sistemas de producción lechera del país. Esta herramienta es el mejoramiento genético por medio de la manipulación molecular o selección asistida por marcadores moleculares (MAS). El objetivo de esta investigación fue determinar la estructura genética de una población Holstein de Antioquia usando el polimorfismo 292 C>T del gen factor del crecimiento insulínico tipo 2 (IGF2a). Una población de 1054 animales de la raza Holstein, distribuidos en 6 subpoblaciones de Antioquia fue evaluada, la genotipificación se llevó a cabo utilizando la técnica PCR-RFLP. Para determinar los estadísticos F de Wright, el equilibrio de Hardy-Weinberg (HW) y las distancias de Nei se utilizaron los programas Genalex 6 y Arlequin 3.5. Las frecuencias alélicas para el polimorfismo 292 C>T de IGF2a fueron para T 0,405 y para C 0,595. La población total se encontró en equilibrio de Hardy-Weinberg, sin embargo, la subpoblación de El Retiro no mostró equilibrio HW. La estructura genética de la población mostró baja diferenciación entre todas las subpoblaciones estudiadas, donde el FST fue 0,002. El nivel de Fit fue de 0,263 y el de Fis de 0,261, ubicándose en los rangos bajos descritos por Wright.
Palabras clave: estadísticos F de Wright, PCR-RFLP, SNPs
Genetic structure of a Holstein population in Antioquia, Colombia, using gene polymorphisms of insulin-like growth factor type 2 (IGF2a)
ABSTRACT
Since the beginning of the field technological development, the livestock production target has been the improvement of the systems through resources optimization to achieve high production levels. Not all of the biological components of such systems, respond the same way to offered nutrients, it is due largely to its genetic component. In response to this situation has emerged an useful tool that could be implemented and applied to daily management of milk production systems in the country. This tool is the genetic improvement through molecular manipulation or molecular assisted selection (MAS) markers. This research target was to determine the genetic structure of a Holstein population in Antioquia by the use of polymorphism 292 C>T from the factor gene of insulin-like growth type 2 (IGF2a). A population of 1054 animals of the Holstein breed, distributed in 6 subpopulations was evaluated in Antioquia. Genotyping was performed using PCR-RFLP technique. The Genalex6 and Arlequin 3.5 software were used to determine the statistical F Wright, the Hardy-Weinberg equilibrium and Nei distances. Allele frequencies for the 292 polymorphism C>T of IGF2a were T (0.405) and C (0.595). The total population was in Hardy – Weinberg balance, however El Retiro subpopulation was unbalanced. The genetic structure of the population showed low differentiation between all studied subpopulations, where the FST was 0.002. In addition, there were found levels of 0.263 Fit and levels of 0.261 Fis rated in the lower ranges described by Wright.
Key words: Wright’s F-statistics, PCR-RFLP, SNPs
Introducción
En Colombia anualmente se producen alrededor de 6717 millones de litros de leche al año, es decir, cerca de 18,4 millones de litros diarios, en dos sistemas de producción de leche, especializada y doble propósito (Federación Colombiana de Ganaderos FEDEGAN, 2015). Los bovinos con los que se cuentan en la actualidad, son el resultado de años de domesticación, en un esfuerzo de productores y mejoradores para lograr animales con características que puedan satisfacer las necesidades de la alimentación humana (López et al., 2011).
Desde tiempos atrás, el sector ganadero ha sido un sector de gran impacto económico, que con el tiempo y con el objetivo de lograr parámetros de cantidad, calidad y sanidad de los productos de origen pecuario, el zootecnista ha hecho del mejoramiento genético cuantitativo una herramienta valiosa, hasta obtener animales con alta producción y calidad, basados en estudios que tienen en cuenta el fenotipo y el medio ambiente en el que se encuentra el animal (Barrientos et al., 2011).
La genética de poblaciones explora los niveles de variación genética dentro y entre las poblaciones que forman a las especies y explica sus patrones en términos de fuerzas evolutivas, además la estructura genética está dada por las diferencias en las frecuencias alélicas de las subpoblaciones que lo conforman, las frecuencias para un gen se pueden ver afectadas por diferentes fuerzas evolutivas como lo son mutación, deriva genética y selección natural, las cuales pueden aumentar la diferenciación genética de las poblaciones. Y lo que es el flujo genético o la migración están encargadas de mantener la homogeneidad genética entre las poblaciones (Eguiarte et al., 2013).
El gen IGF2a (factor del crecimiento insulínico tipo 2) codifica para un factor de crecimiento similar a insulina tipo 2, siendo este expresado en varios tejidos durante el desarrollo de los mamíferos (Hitchins & Moore, 2002). La proteína IGF2, también es conocida como somatomedina A, es un polipéptido que presenta un grado de homología alto con la secuencia de la insulina, desempeña un papel primordial en el crecimiento y desarrollo de los mamíferos, influenciando la división celular, diferenciación, desarrollo de la placenta y una posible regulación metabólica (O’Dell et al., 1997). En los bovinos el gen IGF2a está localizado en el cromosoma 29 (Schmutz et al., 1996; Goodall & Schmutz, 2003), consta de 10 exones, la región codificadora se encuentra en los exones 8, 9 y parcialmente en el exón 10 (Ohlsen et al., 1994; Goodall y Schmutz, 2007).
El objetivo de esta investigación fue determinar la estructura y variabilidad genética de una población de vacas Holstein en Antioquia, Colombia, usando el polimorfismo 292 C>T del gen del factor del crecimiento insulínico tipo 2 (IGF2a).
Materiales y Métodos
Animales y toma de muestras
El estudio se realizó con 1054 animales de raza Holstein, distribuidos en 14 hatos lecheros, ubicados en 6 municipios del departamento de Antioquia: Bello, Belmira, La Unión, Medellín, El Retiro y San Pedro de los Milagros.
Toma de muestra
Para la toma de las muestras de sangre se utilizaron tubos al vacío estériles BD Vacutainer® (Becton, Dickson & Co.), con anticoagulante EDTA (K2EDTA, 1.8 g/ml). con agujas numero 18 (BD Vacutainer®), por duplicado. La muestra de sangre se tomó por punción de la vena coccígea media. Se extrajeron 8ml de sangre en dos tubos Vacutainer (4ml cada tubo), los tubos fueron homogeneizados por inversión, rotulados para su identificación y refrigerados con gel refrigerante durante el transporte hasta el Laboratorio de Ciencias Básicas Animales de la Universidad Nacional de Colombia, sede Medellín, para su respectivo análisis.
Extracción de ADN
El total de la sangre se depositó en un solo tubo, de cada muestra de sangre se extrajo ADN por medio de la técnica “precipitación salina” (Miller et al., 1988). La sangre se centrifugó a 3100 RPM durante 10 minutos, se descartó el sobrenadante conservando el paquete de células en el fondo del tubo; se realizaron lavados por centrifugación usando un buffer de lisis I (10mM Tris HCL pH 7,6, 320 mM de sucrosa, 5 mM MgCI2-6G2O, 1% Tritón X-100), en cada lavado se eliminó el sobrenadante sin perturbar el paquete celular, hasta obtener un botón blanco y sobrenadante incoloro, este paquete de células se resuspendió en 5 ml de buffer de lisis II (10 mM Tris HCL, pH 8.2, 400 mM de NaCl, 2mM de Na2EDTA, proteinasa K (2mg/ml) y 200 ul de SDS). Después de la digestión se adicionó 1.5 ml de solución NaCl saturada, se mezcló por medio de Vortex suave y luego se centrifugó 10 minutos a 2500 rpm en centrifuga refrigerada, se colectó el sobrenadante en un tubo de 15 ml, donde se agregó etanol al 100% a -20ºC, se mezcló suavemente el tubo por inversión hasta que se logró observar la madeja de ADN. La madeja fue resuspendida en etanol al 70% y se centrifugó durante 2 minutos a 2500 rpm en centrifuga refrigerada, se descartó el sobrenadante conservando el ADN en el fondo del tubo; se dejó secar el tubo invertido sobre una toalla de papel y finalmente se resuspendió el ADN en 500 ul de buffer TE 1X pH 8.0 (Tris HCl 1 M y EDTA 0.5M) y se almacenó a 4ºC hasta el momento del análisis (Sambrook y Russell 2001). La cantidad y calidad del ADN extraído se evaluó en gel de agarosa al 0,8%, para la visualización del ADN se utilizó EZ Visión como intercalante y se observó en transiluminador BIO-RAD XR®. La pureza del ADN genómico se determinó en Nanodrop® mediante un análisis de absorbancia en dos longitudes de onda 260/280 solo ADN genómico con una pureza ideal entre 1,8-2,0.
Determinación de las variantes alélicas del gen IGF2A
La genotipificación del sitio 292C>T se llevó a cabo usando reacción en cadena de la polimerasa-polimorfismos de longitud de fragmentos de restricción (PCR-RFLP), con una modificación al método de Goodall & Schmutz (2003), sugerida por Zwierzchowski et al., (2010). Un fragmento de 184 pb, desde el nucleótido 21 al 204 del exón 2 del gen IGF2a bovino (GenBank, Acc. Nº. AY237543), fue amplificado con los oligonucleótidos: IGF2aF: 5`-TTGCCTCCCAGTCAAGCCTG-3`, IGF2aR: 5`-GCTGTGTTGTCTCTGAAGCT-3`
El volumen final de la mezcla para PCR fue de 25 μL, que contenía buffer para PCR a 1X, 15mM de MgCl2, 20mM de dNTP`s, 10mM de oligonucleótidos, 0.8 unidades de Taq polimerasa (Invitrogen®), 50 a 100 ng/μL de ADN genómico, para completar el volumen se usó agua estéril libre de nucleasas.
La PCR se realizó en un termociclador Biometra®, las condiciones de la PCR fueron: desnaturalización inicial 95°C por 5 minutos y luego por 35 ciclos de 1 minuto a 94ºC, 64ºC por 1 minuto, y 72ºC por 1 minuto y una amplificación final de 72°C por 10 minutos. El producto de PCR se visualizó en un gel de agarosa al 2% con EZ visión como intercalante; La caracterización del polimorfismo por RFLP fue realizada sometiendo al fragmento de 184 pb a digestión con 5 unidades de la endonucleasa de restricción BsrI (New England BioLabs, USA), incubada a 65ºC por 3 horas. El producto de la digestión fue visualizado en un gel de agarosa al 3% (GIBCO-BRL, England) con EZ Visión como intercalante. El patrón de restricción obtenido con la enzima BsrI del producto de PCR resulto en tres fragmentos para un animal homocigótico TT 118, 58 y 8 pb; homocigótico CC 184 pb y los animales heterocigóticos CT se obtuvieron los fragmentos de 176, 118, 58 y 8pb.
Análisis estadístico
Frecuencias alélicas y genotípicas
La frecuencia de los alelos se estimó determinando la proporción de cada forma del gen entre el número de copias totales de la población en estudio. Se identificaron los homocigóticos (dos copias del mismo alelo) y los heterocigotos (una copia de cada alelo) y se calculó la frecuencia F de cada alelo contando los homocigotos y añadiendo la mitad de los heterocigotos, con el método descrito por Hartl (2000). Frecuencia total (p) del alelo C en la población es:
P= Fa/a+Fa/b
Frecuencia total (q) del alelo T en la población es:
Q= Fb/b+Fa/b
Dónde: F a/a = homocigótico para CC F a/b= heterocigótico para CT F b/b= homocigótico para TT
Para determinar las frecuencias alélicas y fenotípicas se usó el complemento estadístico GenAlex6 (Peakall y Smouse, 2006).
Diversidad genética La diversidad genética se determinó a través de la heterocigosidad observada (Ho) y la heterocigosidad esperada (He), dentro de las poblaciones y en la población total, utilizando el programa Arlequín v 3,5 (Excoffier et al., 2005).
Estructura genética.
Se determinó el equilibrio de Hardy-Weinberg a través del programa Arlequin 3.5 (Excoffier et al., 2005), con base en las frecuencias alélicas y en la diversidad genética encontrada; en el mismo programa se calcularon los parámetros de estructura poblacional que utilizando el método propuesto por Wright (1989), donde estima el coeficiente de endogamia de una población a partir de subpoblaciones que la conforman. Calculando así el parámetro FIT= HT-Hi/HT, que corresponde a la endogamia total, el parámetro FIS= HS-HI/HS que mide la subdivisión intra poblacional y el FST=FIT-FIS/1-FIS que mide la subdivisión poblacional (Wright 1989); donde HT se refiere a la heterocigosidad esperada en la población total, HI es la heterocigosidad promedio observada en un grupo de poblaciones y HS es la heterocigosidad promedio esperada de cada población. Los cálculos fueron realizados mediante el software Arlequin 3.5 (Excoffier ET AL. 2005), utilizando para ello el análisis de varianza molecular (AMOVA) que permite analizar la variación entre y dentro de poblaciones, con su significancia estadística.
Resultados
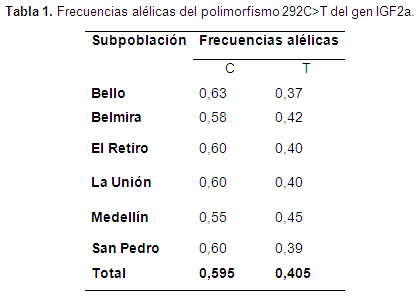
Determinación de las frecuencias alélicas y genotípicas Los fragmentos generados para el polimorfismo 292 C>T del gen del factor de crecimiento insulínico tipo 2 IGF2a que corresponden a un tamaño de 184pb fueron amplificados exitosamente en las 1054 muestras de ADN. La digestión con la enzima de restricción BsrI. permitió caracterizar los genotipos CC, CT y TT, de acuerdo con los tamaños de los fragmentos generados en el total de los individuos analizados. Las frecuencias de los alelos C y T en la población fueron 0,595 y 0,405 respectivamente; la frecuencia de los genotipos CC, CT y TT fue 0,42, 0,35 y 0,23 respectivamente. Las frecuencias alélicas por municipio y la total se presentan en la (tabla 1).
Diversidad genética
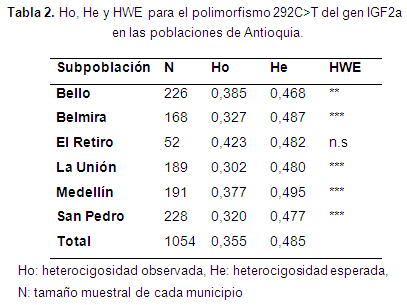
Según las diferencias que se encontraron entre la heterocigosidad observada (Ho) y la heterocigosidad esperada (He), el polimorfismo 292 C>T no se encuentra en equilibrio de Hardy- Weinberg (HWE) en la mayoría de las subpoblaciones, sin embargo, en la subpoblación de El Retiro el polimorfismo 292 C>T presentó equilibrio (p>0,05). Esto puede ser el resultado de los procesos de selección que se han realizado sobre esta raza y puede asociarse a alguna característica de importancia económica.
La heterocigosidad observada y esperada tuvo una variacion de 0,320 a 0,423 y de 0,468 a 0,495 respectivamente.
Estructura genética de las poblaciones
Los estadísticos de Fis, Fit y Fst para la población global fueron 0,261, 0,263 y 0,002 respectivamente, sin diferencias significativas (p<0,05) entre las subpoblaciones.
El parámetro Fis que en la población global fue de 0,261 describe la distribución de los genotipos encontrados en las subpoblaciones, lo que indica que el valor encontrado se encuentra en las proporciones esperadas cuando los apareamientos son estrictamente al azar, además donde se observó la presencia de homocigosis.
El valor de Fit presentó un valor de 0,263, con tendencia hacia la endogamia; sin embargo, los valores de Fis no fueron significativos (p>0,05), que no permiten ser categóricos al expresar esta tendencia en la población total de Antioquia.
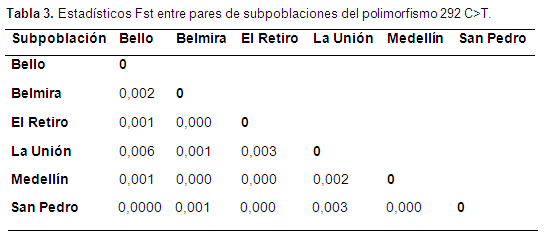
Los Fst para la población total mostraron un valor de 0,002, que muestra una baja o nula diferenciación genética entre las subpoblaciones (tabla 3), que confirman una baja magnitud, según los rangos descritos por Wright (1989); que muestran una estructura genética dentro de estas poblaciones, y no es posible analizar Antioquia como una sola unidad, aunque sea muy baja la magnitud. Los parámetros estuvieron entre 0,000 y 0,006, encontrándose todos según los rangos propuestos por Wright en la categoría de baja magnitud (Wright, 1969).
Los estadísticos de Wright miden la subdivision de las poblaciones y determinan si existen o no diferencias en las frecuencias alélicas de las 6 subpoblaciones, todas las subpoblaciones estudiadas se encontraban en equilibrio de HE, excepto la subpoblación de El Retiro, esto puede atribuirse a que al ser las subpoblaciones municipios dedicados a la lechería especializada, hubo aumento de selección por parte de los ganaderos, que ocasionó el desequilibrio y favoreció más un genotipo que otro; sin embargo, para que una población se considere en equilibrio de HW debe de cumplir los supuestos: ser una población grande, tener presente entrecruzamiento al azar, no presentar un cambio alélico, no migración y que no exista selección.
Discusión
Las frecuencias alélicas obtenidas del gen IGF2a en este estudio a partir del polimorfismo 292 C>T fueron similares respecto a estudios realizados en otras partes del mundo en vacas Holstein, donde el polimorfismo del alelo C presentó mayor frecuencia respecto al alelo T (Goodall & Schmutz, 2003; Bagnicka et al., 2009; Zwierzchowski et al., 2010), caso contrario a estudios realizados en ganado Hereford, Simmental, Charolais y Angus, donde la frecuencias del alelo T es menor a las presentadas por otros autores. Sherman et al. (2008) reportó una frecuencia para el alelo T de 0,19 para la raza Angus y 0,16 en raza Charolais.
El valor mayor de la frecuencia para el alelo C fue que el aleo T; son similares a los resultados de estudios realizados en Holstein, donde se encontró que el alelo C es más frecuente que el T (Goodall y Schmutz, 2007; Sherman et al., 2008 ; Bagnicka et al., 2009; Zwierzchowski et al., 2010,). En relación, se encontró el genotipo CC como el genotipo más frecuente, el resultado es similar a lo obtenido por Bagnicka et al. (2009) (CC 0,49), mientras que en lo reportado por Zwierzchowski et al. (2010) en Holstein polaco (CC 0,25), y el genotipo heterocigoto fue el más frecuente (CT 0,48).
Las poblaciones evaluadas en Antioquia no presentaron desviaciones significativas (p>0,05) del equilibrio de Hardy- Weinberg para el gen IGF2a, lo que indica que las 6 subpoblaciones que la conforman no han sido sometidas a procesos de selección intenso sobre este gen, sin embargo la subpoblacion de El Retiro presentó un desviación significativa en el equilibrio HW.
Diferentes autores han investigado sobre la estructura genética de la población del ganado Holstein en Colombia, a través de la evaluación de varios polimorfismos de importancia económica, genes como lactoferrina (LTF), hormona del crecimiento bovina (bGH), prolactina (PRL) (Rincón et al., 2013; Rodríguez et al., 2013). Para estos genes las poblaciones se han encontrado en HW.
La estructura genética observada en este estudio para el gen IGF2a a partir de los estadísticos de F de Wright, obtuvo valores en un rango bajos para Fis: 0,261 no significativos, por lo que no es posible decir que existen heterocigosis ni homocigosis dentro del total de la población de Antioquia. En estudios de Rodríguez et al. (2013), se encontraron valores Fis en un rango medios a partir del gen LTF, resultados similares a Rincón et al. (2013), que observó valores medios para Fis pero en el gen de bGH.
Los Fst para la población total evaluada presentaron una variación de baja magnitud Fst: 0,002, que no presentó significancia, por lo que puede indicar una baja o nula diferenciación genética entre las subpoblaciones investigadas; por lo tanto, la población Holstein de Antioquia se comporta como una única población.
Este estudio es un antecedente para determinar cuáles genes son los que presentan mayor relación con parametros productivos de importancia. El gen IGF2a se ve involucrado en el desarrollo mamario, ademas juega un papel muy importante en la diferenciaciòn y crecimiento muscular y se ha relacionado con características de importancia económica en otros países, por lo cual se puede convertir en un candidato para marcador en el uso de la selección asistida MAS.
Referencias bibliográficas
Cómo citar: García-Valencia, M.; López-Herrera, A.; Echeverry-Zuluaga, J. Estructura genética de una población de bovinos Holstein en el departamento de Antioquia, usando polimorfismos del gen del factor de crecimiento insulínico tipo 2 (IGF2a). Revista Veterinaria y Zootecnia, v. 11, n. 1, p. XX-XX. DOI: 10.17151/vetzo.2017.11.1.2 |







 | Today | 507 |
 | Yesterday | 631 |
 | This week | 4397 |
 | Last week | 4842 |
 | This month | 15280 |
 | Last month | 15809 |
 | All days | 1359347 |
Online Now: 8
Your IP: 3.147.86.143
MOZILLA 5.0,
Today: November 23, 2024
Your IP: 3.147.86.143
MOZILLA 5.0,
Today: November 23, 2024